Editor's Note. This essay was subsequently published as Dillon, R.T., Jr. (2019b) Everything changed when I met Samantha. Pp 67-72 in The Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
Three years ago I posted a series of five essays on this blog entitled, “The Lymnaeidae 2012” [1-5]. My primary motivation was the imminent expansion of the FWGNA project into the Mid-Atlantic States, where I expected that we would come into contact with the range of an enigmatic group of lymnaeids called the “stagnicolines.” And a couple comprehensive molecular phylogenetic studies had also recently been published that I thought might cast some light on systematic relationships in the group.
Three years ago I posted a series of five essays on this blog entitled, “The Lymnaeidae 2012” [1-5]. My primary motivation was the imminent expansion of the FWGNA project into the Mid-Atlantic States, where I expected that we would come into contact with the range of an enigmatic group of lymnaeids called the “stagnicolines.” And a couple comprehensive molecular phylogenetic studies had also recently been published that I thought might cast some light on systematic relationships in the group.
 |
| Samantha |
Who was Ms. Samantha Flowers? And where has she gone, long time
passing? Our good friend Tom Duda from
the University of Michigan introduced her at the AMS meeting in Cherry Hill in
June of 2012, and we kept in touch until August of 2013, at which point she
disappeared. But she left behind a
tangled body of potentially important research on the genetics of the stagnicolines,
which we will sort through together, as this, our fresh series of essays on the
Lymnaeidae unfolds.
So when last we left our story, it may be recalled
that the Baker/Burch system for the classification of the North American
Lymnaeidae recognizes 21 species of stagnicoline lymnaeids in two subgroups. The dark-bodied populations of bogs, marshes
and vernal ponds bearing slender shells include elodes (Say 1821), exilis (Lea
1834) and three others more recently described.
The pale-bodied inhabitants of open waters, bearing broader, more robust
shells include catascopium (Say 1816), emarginata (Say 1821) and 14 others more
recently described.
Half of the challenge with which we wrestled in
2012 was the relationship between our New World stagnicolines and those of the
Old. In 1951, my hero Bengt Hubendick
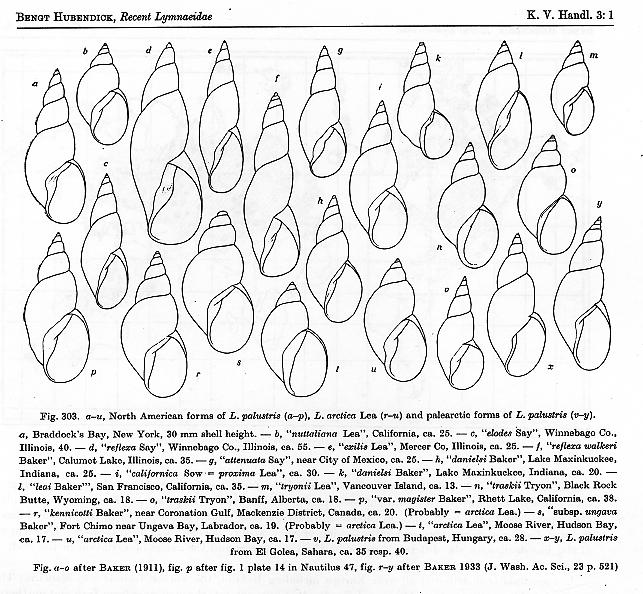
synonymized all the dark/skinny species of North America under the European
palustris (Muller 1774). Hubendick’s
figure of L. palustris is reproduced below – click for a full-sized version,
with caption [6]. But by the 1960s evidence
had begun to accumulate that the European palustris is a complex of several
cryptic species, distinguishable only by detail of reproductive anatomy. And in my essay of 10May12 [2], I offered
evidence that at least two cryptic species of dark/skinny stagnicolines also seem
to inhabit the ephemeral ponds and marshes of NW Pennsylvania.
Although I did not mention it at the time, shortly
after I published my “cryptic stagnicoline” essay of 10May12, I asked my good
friends Kip Brady and Andy Turner to send samples of their enigmatic Pennsylvania
populations to Charleston, which I dissected, comparing details of their reproductive
anatomy to figures from the European literature. Alas, I was unable to distinguish any of
these populations anatomically, and let the matter drop. But the cryptic stagnicolines of Brady &
Turner turned out to be key to disentangling Samantha Flowers’ research
results, when we were finally able to examine them in 2015. So keep this in the back of your mind.
The other half of the challenge to working out the
systematic relationships among our American stagnicolines is their great potential
for ecophenotypic plasticity of shell.
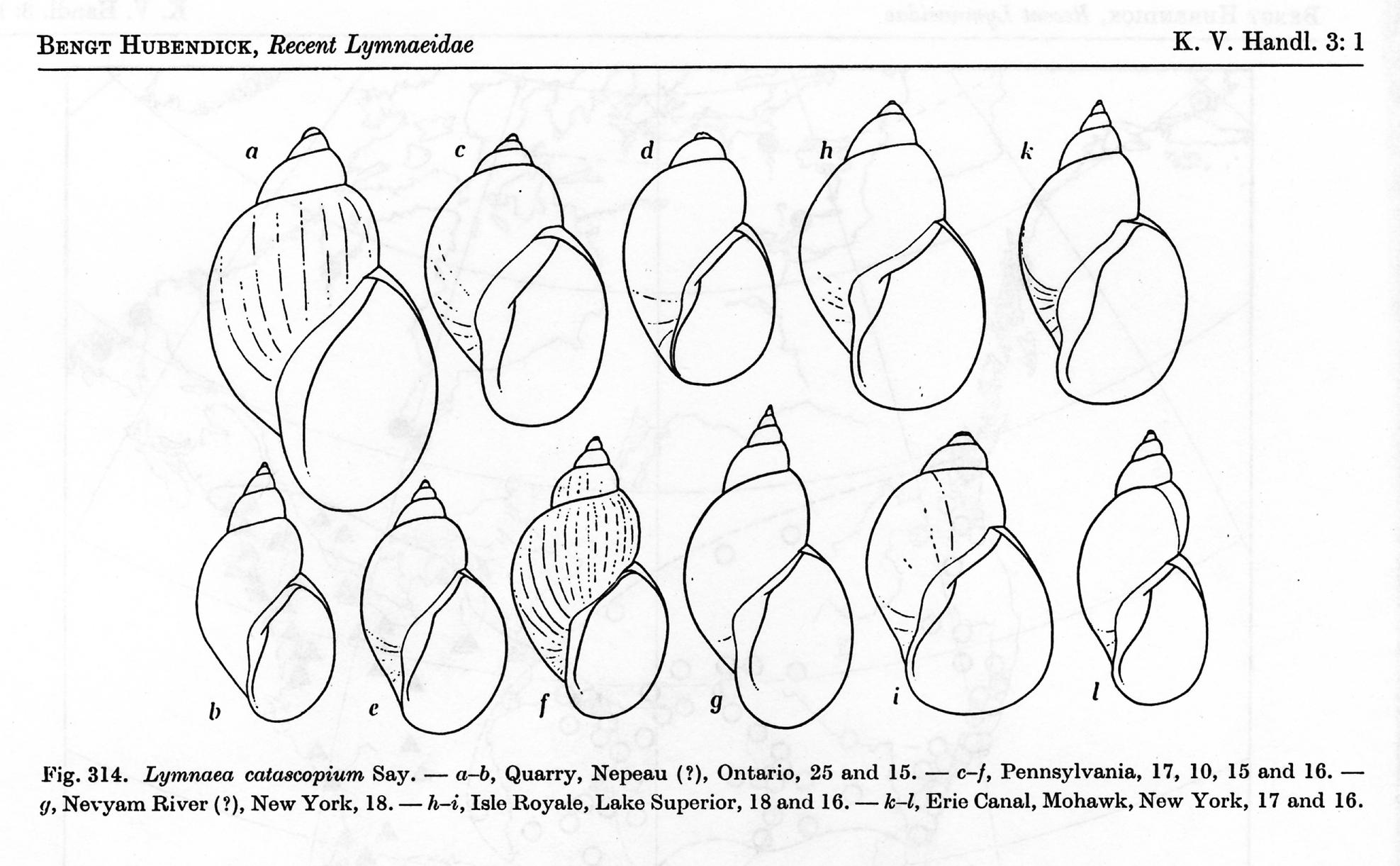
Given the large body of research results such as those of Christer
Bronmark on European Lymnaea peregra (aka “Radix balthica”), it is not
inconceivable that the robust shells with enlarged body whorls born by
populations we call catascopium or emarginata here in North America arise as an
ecophenotypic response to life on solid substrates in open waters, exposed to
fish predation [4]. There may be no
additively heritable basis for the distinction between the broad, heavy shells
of the catascopium/emarginata subgroup and the slender, gracile shells of the elodes/exilis
subgroup whatsoever. Hubendick’s figure
of L. catascopium is reproduced below – click for a full sized version, with
caption.
So with the potential for cryptic speciation and shell
ecophenotypic plasticity firmly in mind, in the spring of 2012 I designed a genetic
survey of the North American stagnicolines.
My plan was to sample populations from the type localities of the four
oldest nomina – catascopium, emarginata, elodes, and reflexa. And along with each topotypic population, I
also hoped to sample a nearby population bearing the opposite shell form. My hypothesis was that each broad, heavy
open-water population would prove most genetically similar to its local
slender/gracile marsh-dwelling population.
For example, the type locality of L. catascopium
is the Delaware River at Philadelphia, and the type locality of L. emarginata
is “Lakes of Maine.” In 2012 it seemed
likely to me that Delaware River catascopium might prove most genetically
similar to the populations of (nominal) L. elodes that I expected to find in
the marshes of Delaware tributaries in eastern Pennsylvania, and that Maine emarginata
might prove most genetically similar to populations of nominal L. elodes
sampled from the marshes and vernal ponds of Maine. I imagine my readership will recognize this
study design as the same I have used to confirm “cryptic phenotypic plasticity”
in a variety of pleurocerid taxa in recent years [7].
And so I mapped out an itinerary for my 2012 field
season. I set aside six days in June for what (I presciently imagined)
might be a challenging quest to re-discover L. catascopium in the Delaware
River, and L. elodes in vernal habitats of the Delaware Valley, after which I
planned to attend the meeting of the American Malacological Society,
conveniently scheduled in the Philadelphia suburb of Cherry Hill (NJ) June 16 –
21.
Everything changed when I met Samantha. But coming next month… “The type locality of
Lymnaea catascopium.”
Notes
[5] The Lymnaeidae 2012: Fossarine Football [7Aug12]
[6] Hubendick, B. (1951) Recent Lymnaeidae, Their Variation, morphology, taxonomy, nomenclature and distribution. Kungl. Svenska Vetenskapskademiens Handlingar. Fjarde Serien Band 3, No. 1. Stockholm: Almquist & Wiksells.
See The Classification of the Lymnaeidae [28Dec06]
[6] Hubendick, B. (1951) Recent Lymnaeidae, Their Variation, morphology, taxonomy, nomenclature and distribution. Kungl. Svenska Vetenskapskademiens Handlingar. Fjarde Serien Band 3, No. 1. Stockholm: Almquist & Wiksells.
See The Classification of the Lymnaeidae [28Dec06]





