Editor’s Note – This essay was subsequently published as: Dillon, R.T., Jr. (2023c) Albinism and Sex Allocation in Physa. Pp 169 – 175 in The Freshwater Gastropods of North America Volume 7, Collected in Turn One, and Other Essays. FWGNA Project, Charleston, SC.
Last month we reviewed, in personal and anecdotal fashion,
the events leading up to (what has turned out to be) a thirty-year research
program on the North American Physidae [1].
And when we left the story, in the summer of 1989, the entire second
floor of the College of Charleston Science Center was in danger of being overwashed
by a sea of 10 oz disposable cups of snails.
On July 25, 1989, our good friend and former student Amy
Wethington wrote this in her research notebook: “Found albinos (or what at least
thought was albinos) in the following cups.”
She then went on to list six culture vessels, some prefaced by the
number 15, others prefaced by the numbers 27 and 29.
These numbers corresponded to isofemale lines. The culture vessel labelled "15-06," for example, was the sixth sibship laid by female number 15, of the 30 we had isolated for our sperm storage experiment. And inside culture vessel 15-06, Amy had found at least one snail that was entirely unpigmented, body and shell, with
unpigmented eyes. All were juveniles or subadults at the point of their identification. Amy carefull lifted each of the albino snails out of the cups in which they were born and separated them by line.
Amy’s discovery opened
an entirely new research direction for us, which extended over ten years and
ultimately yielded her MS degree from The College and six papers in
peer-reviewed journals. These papers
were not directly relevant to the question that had nagged me since my high
school days, the actual identity of the snails now covering every square inch
of benchtop in the College of Charleston genetics lab. But they armed Amy and me with important
insights into the biology of our study organism – insights that would become vital
as our research program unfolded.
Pop quiz everybody!
What is a complementation test?
Most of you probably heard that term in your undergraduate genetics
class, and figured you’d never need it again.
A complementation test is a genetic cross conducted to
determine whether phenotypic variants are allelic. So for example, Amy had discovered albinos in
three different lines of Physa, each founded by single isolated females
collected some weeks previous from a neighborhood pond – Number 15, Number 27,
and Number 29. In late August, she found
albinos in isofemale line Number 7 as well.
Are all of these snails albinistic for the same reason? In other words, do they all have mutations at
the same step in the biochemical pathway leading to pigmentation? Then the best hypothesis would be that they
are all “allelic,” homozygous recessive for alleles at the same locus. Or might they have mutations at different
steps in the pigment pathway? In which
case they are non-allelic; more than one locus is involved.
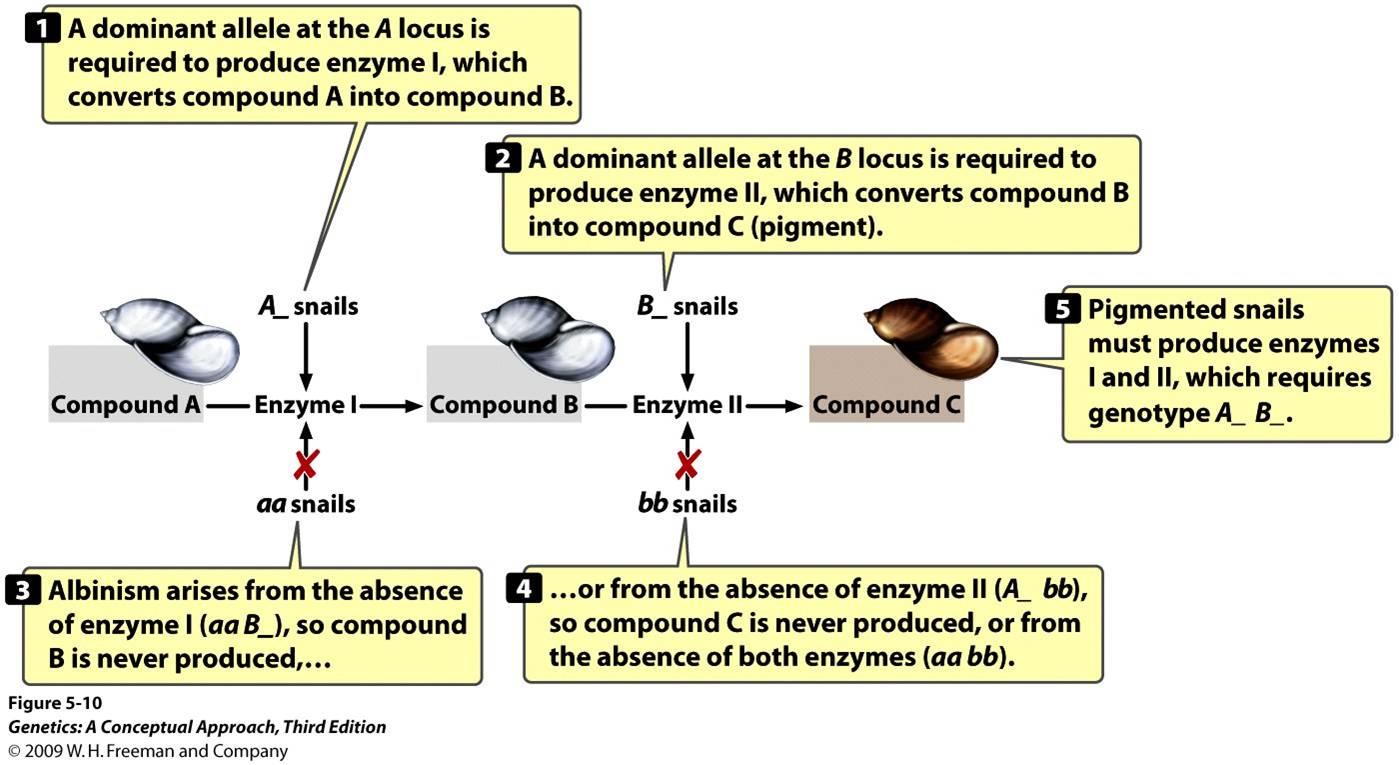
And here’s another little dose of your old genetics
professor kicking in. During the second week of my Junior-level Genetics class,
we review the phenomenon of epistasis – where one locus masks a second. When Amy and I intercrossed the F1 offspring from
our 7 x 15, 7 x 27, or 7 x 29 hybrid lines, we obtained a 9:7 ratio of
pigmented to unpigmented F2, the classic signature of reciprocal recessive
epistasis. Amy and I published that
paper in the Journal of Heredity in 1992 [2].
I’ll never forget the morning in the fall of 2002, ten years
later, when a colleague brought a copy of the brand new textbook by B. A. Pierce,
Genetics: A Conceptual Approach, into my office. Pierce had selected our results on albinism
in Physa to illustrate the concept of epistasis. Just to see the diagram reproduced above, reproduced
as Figure 5.10 in a genetics textbook used by thousands of college students,
was the biggest thrill I have ever enjoyed with my pants on.
Amy and I used the albinism genes as tools to study a broad
swath of reproductive biology in Physa, ultimately yielding the papers
footnoted [2] through [7] below. The
most interesting study, I think, is the one on protandric hermaphroditism which
we published in the Proceedings of the Royal Society in 1993 [3]. The experimental setup was as clever as I can
get.
Imagine two individual Physa fixed for albinism at different
loci – call one “7-experiment” and the other “29-challenge.” The 7-experiment snail is a mature adult; he has
been reared in isolation, and is now self-fertilizing, yielding (of course)
albino progeny. The 29-challenge snail
is a hatchling… maybe just a couple weeks old – still very juvenile.
Introduce the 29-challenge snail into the cup with the 7-experiment
snail for one day, and then separate them again. Initially, no change is expected. The 7-experiment snail should continue to lay
self-fertilized eggs, and the 29-challenge snail lays no eggs at all. Then do that again, at week two. And do that again, at week three. And so forth.
When the 7-experiment snail begins to lay pigmented eggs, we know that
the 7-experiment snail has matured as a female.
And when the 29-challenge snail begins to lay pigmented eggs, we know
that the 7-experiment snail has matured as a male.

The experiments that Amy and I conducted according to this
design ultimately showed that Physa are “protandric hermaphrodites.” Male reproductive maturity was reached at
mean age of 5.7 weeks in our culture conditions, with female reproductive
maturity added at a mean age of 7.3 weeks.
And reared in isolation, our Physa delayed self fertilization to a mean
age of 22 weeks; certainly at a significant cost to fitness in the wild.
At our sample sizes we were also able to detect low
frequencies of all sorts of wonderful reproductive variance, including
autosterility, outcross male-sterility, outcross female-sterility, and outcross
double-sterility. Although most snails
matured first as male and then as female, we discovered a few cases of
simultaneous development, a couple female-first snails, and some snails that
passed through brief periods of self-fertilization before outcrossing as
females. It’s amazing that anybody knows
this. Or cares, come to think about it.
We completed several other interesting studies involving
those complementing albino lines from 1990 to 1992, when Amy graduated and left
Charleston - our (1996) study of gender choice and gender conflict (Amy’s MS
thesis, footnote 5) and our (1997) estimates of lifetime fitness and inbreeding
depression [6]. And we still had at
least one or two cool experiments mapped out on the Genetics Lab
chalkboard. But alas. After a couple years in culture, all our
albino lines seemed to quit outcrossing.
And by the mid-1990s, they had died out, victims of inbreeding
depression, I feel sure.
But a couple nice young undergraduate students and I
published one last albino paper in 2005 [7], which I still remember with considerable
fondness. Walking along the edge of our
pond at Charles Towne Landing in September of 2002, I happened to discover an adult
albino Physa – the first I had ever seen in many years of observation. Assuming (what must be) a very low background
frequency of albinism in the wild population, it is reasonable to guess that that
this single albino snail had already been wild-inseminated by a homozygous
pigmented partner.
All the experiments we had done in the previous 10 – 12
years had involved lab lines in culture.
So Tommy McCullough and Charles Earnhardt and I isolated this single
snail, whom the students named “White Fang,” in a 10 oz cup [8]. She laid 109 eggs over the next week, which
hatched into 30 viable pigmented offspring, and 6 albinos. We advanced her to a second cup and got
similar results. And so forth, on until
February of 2003, when White Fang finally went up to Fort Yukon.
Totaled over our 20 weeks of observation, White Fang laid
1,566 eggs, which hatched into 550 viable hatchlings, of which 35 were albinos,
for an effective self-fertilization rate of 6.4%. This is the best estimate of self-fertilization
in any (mixed-mating) population of pulmonate snails of which I am aware [9].

And as of week 20, White Fang was still laying pigmented and
albino eggs at the same frequency that she was producing them at week 1. Finally we had answered the sperm-storage
question that had prompted this entire research program, back when Amy and I got
our start in 1989. Arbitrary adult Physa
sampled from wild populations apparently do not ever run out of allosperm, even
if isolated. They are inseminated for life.
Well, by 2005, our Physa project had diverged in all sorts
of fresh directions. Which brings us
back to the main theme of this series of essays.
Notice that in all those papers Amy and I published using
our albino lines in the 1990s, we were still referring to our study organism as
“Physa heterostropha pomilia.” But by
2005, White Fang was identified as an individual “Physa acuta.” What happened? To be continued!
Notes
[2] Dillon, R.T. and A.R. Wethington (1992) The inheritance
of albinism in a freshwater snail, Physa heterostropha. Journal of Heredity
83:208-210. [PDF]
[3] Wethington, A.R., and R.T. Dillon (1993) Reproductive
development in the hermaphroditic freshwater snail, Physa, monitored with
complementing albino lines. Proceedings of the Royal Society (London) B
252:109-114. [PDF]
[4] Dillon, R.T., and A.R. Wethington (1994) Inheritance at
five loci in the freshwater snail, Physa heterostropha. Biochemical Genetics
32:75-82. [PDF]
[5] Wethington, A.R., and R.T. Dillon (1996) Gender choice
and gender conflict in a non-reciprocally mating simultaneous hermaphrodite,
the freshwater snail, Physa. Animal Behaviour 51: 1107-1118. [PDF]
[6] Wethington, A.R., and R. T. Dillon. (1997) Selfing,
outcrossing, and mixed mating in the freshwater snail Physa heterostropha:
lifetime fitness and inbreeding depression. Invertebrate Biology 116:
192-199. [PDF]
[7] Dillon, R. T., T. E. McCullough, and C. E. Earnhardt.
(2005) Estimates of natural allosperm storage capacity and self-fertilization
rate in the hermaphroditic freshwater pulmonate snail, Physa acuta. Invertebrate Reproduction and Development 47:
111-115. [PDF]
[8] Had one of the undergraduates recently read Jack
London’s (1906) novel? I don’t know. But I am pretty sure that the name “White
Fang” was supposed to be ironic.
[9] Yes, two undergraduates and I got a paper published in an
international journal with 20 plastic cups, a teaspoon of fish food, and one,
single snail. You really don’t need big
grants to do good science. You don’t
need any money at all. That’s what I
have always told myself, pretty much every year over my entire career, as my
grant proposals have been rejected.