Back in July of 2008 (1) we reviewed the relationship between gene trees and species trees, a subject that has become fashionable at the highest levels of evolutionary science. The phenomenon that leads to differences between the two types of evolutionary tree is usually called “lineage sorting” by phylogenetic systematists, who otherwise ignore it, hoping it will go away. But population geneticists, who tend to run their gene trees backward and call the phenomenon "coalescence," have pointed out that the differences in a set of gene trees can be used to date divergence events in a species tree. This may be the one thing that gene trees are actually good for.
But what is a "species tree?" Wayne Maddison, in his seminal 1997 paper on the subject (2), had in mind a phylogeny of bona fide biological species, "when reproductive communities are split." But reproductive isolation need not evolve between a pair of populations for lineage sorting to commence in their gene pools. The clock starts ticking on the gene trees as gene flow becomes disrupted for any reason between any pair of populations, reproductively isolated or not. So although the scientific community most actively involved in this area of research has always used the term "species tree" to describe the phylogeny they are comparing to their gene trees (3), the term "population tree" would clearly be much more accurate.
Genuine species trees turn out to be surprisingly difficult even to visualize, much less to work out. I did not appreciate the difficulty myself until I tried to draw one, or actually a set of them, for the paper Amy Wethington, Chuck Lydeard, and I recently published in BMC Evolutionary Biology (4).
Our new paper summarizes over ten years of research we've conducted on the evolution of reproductive isolation in Physa, including papers we've published comparing Charleston P. acuta, P. gyrina, P. pomilia, and P. carolinae (5). Brand new for 2011 we've added a second population of P. acuta, sampled from Philadelphia.
Here's an obvious attribute of the word "species" that I think we may all take too much for granted. The word "species" is relational, like the word "brother." It is not a point-character that can be measured on an OTU and cast simply on a phylogenetic tree. So when Amy, Chuck, and I undertook to add a second population of P. acuta to the species tree we had been building for years, we added a row to our triangular matrix. Between acuta, carolinae, pomilia and gyrina there are 3+2+1=6 sets of pairwise mate choice tests to be completed, and 6 corresponding sets of no-choice hybridization experiments. Adding the Philadelphia acuta population added 4 more sets of both, almost doubling our effort. A pain in the butt.
And here’s another bullet point we might profitably highlight before drawing our first species tree. The single, original population at the base of our hypothetical tree was (of course) reproductively compatible with itself. What evolves, as a species tree splits, is reproductive incompatibility. So (to borrow a term from cladism, which I hate) retention of reproductive compatibility is a "symplesiomorphy." I can still remember the day back in the early 1980s when an ANSP ichthyologist first called my attention to a paper by Donn Rosen rejecting the biological species concept because an ideologically pure classification cannot be based on symplesiomorphy (6). I'm still pissed off about that.
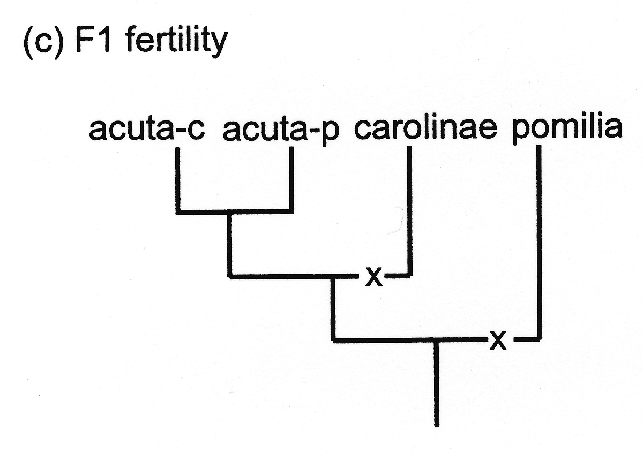 In any case, I should think that most of us, if we were to visualize a bona fide species tree, would begin with something like tree (c) at left for F1 fertility in Physa. No hybrids were recovered from any of the no-choice experiments testing gyrina against any of the other four populations, so gyrina is not shown in tree (c). At least some hybrids were born in the other 3+2+1 experiments, the ones between Charleston acuta-c and Philadelphia acuta-p proving perfectly fertile, the other 3+2 categories of hybrids not. So starting with hybrid fertility at the base of the tree, the situation may have been as simple as two separate mutations evolving at a single locus, from fertile to sterile. The actual phenomenon in Physa is almost certainly controlled by multiple loci, but our observations can be modeled very simply, as just the one.
In any case, I should think that most of us, if we were to visualize a bona fide species tree, would begin with something like tree (c) at left for F1 fertility in Physa. No hybrids were recovered from any of the no-choice experiments testing gyrina against any of the other four populations, so gyrina is not shown in tree (c). At least some hybrids were born in the other 3+2+1 experiments, the ones between Charleston acuta-c and Philadelphia acuta-p proving perfectly fertile, the other 3+2 categories of hybrids not. So starting with hybrid fertility at the base of the tree, the situation may have been as simple as two separate mutations evolving at a single locus, from fertile to sterile. The actual phenomenon in Physa is almost certainly controlled by multiple loci, but our observations can be modeled very simply, as just the one.The situation cannot be so simple for hybridization, however. Of course the Charleston acuta-c and the
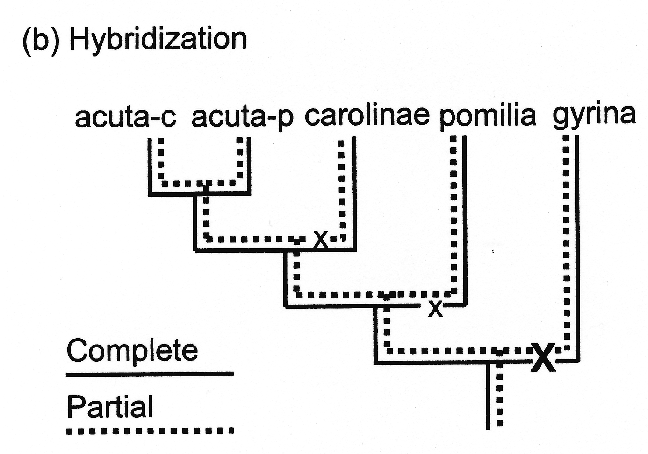 Philadelphia acuta-p will hybridize, and as we just noted, gyrina does not hybridize in our experimental conditions with any of the other species at all (7). Our Physa carolinae population freely hybridizes with either population of acuta, while P. pomilia partially hybridizes with either population of acuta, yielding mixtures of selfed and outcrossed F1 progeny. Our carolinae x pomilia no-choice experiments have not yielded any hybrids.
Philadelphia acuta-p will hybridize, and as we just noted, gyrina does not hybridize in our experimental conditions with any of the other species at all (7). Our Physa carolinae population freely hybridizes with either population of acuta, while P. pomilia partially hybridizes with either population of acuta, yielding mixtures of selfed and outcrossed F1 progeny. Our carolinae x pomilia no-choice experiments have not yielded any hybrids.My best effort to depict this messy set of relationships is shown in tree (b) at right above. Our model suggests two loci, a "complete" locus J and a "partial" locus K, at which unique compatibility alleles segregate, locus J epistatic over locus K. See the text of our paper for the details. The bottom line is, however, that the simplest model I can conceive of for the evolution of barriers to hybridization in Physa is not especially simple.
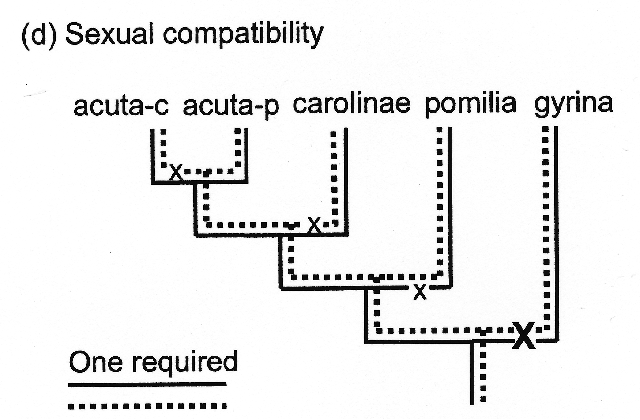 Nor is the situation on the evolution of sexual incompatibility simple, at all. Although our mate choice tests did show prezygotic reproductive isolation between P. carolinae and Charleston acuta-c, no behavioral barriers seem to be in place to lower the frequency of copulation between carolinae and Philadelphia acuta-p. Those observations seem to require another two-locus model (tree d), again see our paper for the gory details.
Nor is the situation on the evolution of sexual incompatibility simple, at all. Although our mate choice tests did show prezygotic reproductive isolation between P. carolinae and Charleston acuta-c, no behavioral barriers seem to be in place to lower the frequency of copulation between carolinae and Philadelphia acuta-p. Those observations seem to require another two-locus model (tree d), again see our paper for the gory details.Our paper has a very mild conclusion, given the years of sturm und drang through which we passed to arrive upon it. The three species trees shown above, with their minimum of five genes for reproductive isolation as postulated, do indeed match the CO1+16s mtDNA gene trees previously published by Wethington & Lydeard and Wethington et al (8). I was kind-of hoping that they would not, because our conclusions would have been cooler. But (try as I might) I cannot rearrange the three species trees shown above to make them any simpler, to not match the gene tree.
I'm sure this isn't the first time in the history of science that the relationship between gene trees and bona fide species trees has ever been tested, but I don't know of any other. And the match seems to be a good one, darn it. But I am not issuing a warrant to all you gene tree jocks out there to get cocky.
Notes
(1) Gene Trees and Species Trees [15July08]
(2) Maddison, W. 1997. Gene trees in species trees. Systematic Biology, 46, 523-536.
(3) For example see Hudson, R. R. (1992) Gene trees, species trees, and the segregation of ancestral alleles. Genetics 131: 509-512. Wakeley, J. (2008) Coalescent Theory, An Introduction. Roberts & Co., Greenwood Village, CO 326 pp. Degnan et al. (2009) Properties of consensus methods for inferring species trees from gene trees. Syst. Biol. 58: 35-54.
(4) Dillon, R. T., Jr., A. R. Wethington & C. Lydeard (2011) The evolution of reproductive isolation in a simultaneous hermaphrodite, the freshwater snail Physa. BMC Evolutionary Biology 11:114.
Online clickable version [html]
Standard [pdf]
(5) Dillon, R.T., Jr., Robinson, J. & Wethington, A. 2007. Empirical estimates of reproductive isolation among the freshwater pulmonate snails Physa acuta, P. pomilia, and P. hendersoni. Malacologia, 49, 283-292. [pdf] Dillon, R. T., Jr., Earnhardt, C. & Smith, T. 2004. Reproductive isolation between Physa acuta and Physa gyrina in joint culture. American Malacological Bulletin, 19, 63-68. [pdf] Dillon, R. T., Jr. 2009. Empirical estimates of reproductive isolation among the Physa species of South Carolina (Pulmonata: Basommatophora). The Nautilus, 123, 276-281. [pdf] That last paper was featured in my blog post entitled "True Confessions: I described a new species" [7Apr10]
(6) Rosen, D. E. (1979) Fishes from the uplands and intermontane basins of Guatemala: revisionary studies and comparative geography. Bull. Amer. Mus. Nat. Hist. 162: 270-375.
(7) Although we recovered no hybrids from our no-choice experiments published in 2004, Tom Smith and I did discover a couple acuta x gyrina hybrids naturally occurring on the margins of the Delaware River at Washington Crossing. They showed up quite unexpectedly on allozyme gels. That's all I know, until God grants me another life, or a few more good students.
(8) Wethington AR, Lydeard C (2007) A molecular phylogeny of Physidae (Gastropoda: Basommatophora) based on mitochondrial DNA sequences. J Moll Stud 73: 241-257. [12Oct07]Wethington AR, Wise J, Dillon RT Jr (2009) Genetic and morphological characterization of the Physidae of South Carolina (Pulmonata: Basommatophora), with description of a new species. Nautilus 123: 282-292. [pdf]






{kind=link}