Editor’s Note – This essay was subsequently published as:
Dillon, R.T., Jr. (2023b) On the Trail
of Professor Troost. Pp 35 – 40 in The
Freshwater Gastropods of North America Volume 6, Yankees at The Gap, and Other
Essays. FWGNA Project, Charleston, SC.
The Dutch-American geologist Gerard Troost [1] had already
led a full life when he stepped off the boat in Philadelphia in 1810. He had earned a doctorate in medicine from
the University of Leyden and done graduate work in crystallography at
Paris. He had been wounded in the
Napoleonic Wars, served as chief scientist on an expedition to Java, and was
captured and ransomed by privateers, twice, both French and English. Sometimes we imagine that our lives today are
interesting.
 |
| Gerard Troost (1776-1850) |
In 1812 Troost was elected the first president of the
Academy of Natural Sciences, a post he held for five years, tutoring Isaac Lea
and Lardner Vanuxem. Then in 1825 he
sailed down The Ohio to “happiness, enlightenment and prosperity” at New
Harmony, Indiana, with Thomas Say at his side.
Gerard Troost was not a malacologist, but he certainly had the names of a
couple good ones in his Rolodex.
Troost was called away from New Harmony after just two years
singing in the choir utopian by the offer of a professorship at the University
of Nashville, from whence he was appointed the Tennessee State Geologist in
1831. These duties sent him on lengthy explorations
throughout the Volunteer State, bringing him to the verge of many rivers and
streams blessed with rich faunas of freshwater mollusks. He was often accompanied in the field by Dr.
Richard Owen Currey (1816 – 1865), whom we mentioned in October [2], who
assumed Troost’s duties as Professor of Geology in Nashville when Troost died
in 1850.
Troost apparently began sending shells to his good friend
Isaac Lea in Philadelphia very shortly after his 1831 appointment and continued
for quite a few years. Leafing through
the Lea bibliography in Scudder [3] I see that at least 25 – 30 new species of
unionids Lea described in the 1830s and 1840s were “sent to me by Professor
Troost.” And it was in honor of
Professor Troost that Lea described Melania troostiana around 1838ish,
approximately.
It will be remembered from last month’s post that through
most of his career Isaac Lea was locked in a torrid race for the naming of
species, and that the precise dates of his publications, calculated down to the
afternoon, mattered a great deal, at least to him. To establish what any of those dates might
actually have been, however, demands scholarship of a higher caliber than the
popgun your humble essayist comes packing.
Lea read his initial description of Melania troostiana in
brief, Latinate form before the American Philosophical Society on November 4,
1836. His (more complete, English)
description appeared in the Transactions Volume 6 (New Series), Article 1, which
(according to Scudder) was “printed and ready for publication” June 15, 1838 [4]. If you download a copy of Transactions Volume
6 from the Biodiversity Heritage Library, however, it very clearly states 1839
on the title page. And (apparently in
error) both Tryon [5] and Goodrich [6] give Lea’s date for the publication of
troostiana as 1841. Burch [7] and Graf [8]
harken back to 1838.
But here is the thing that matters to us today. Melania troostiana was early. Regardless of its actual publication date, Lea’s
description of Gerard Troost’s eponymous pleurocerid certainly preceded the
torrent of gastropod nomina that spilled from his pen beginning in 1841 with
his “New Fresh Water and Land Shells” series.
If you run your finger down the list of 199 canonical
pleurocerid nomina forwarded to us by Goodrich/Burch, troostiana falls out #39
[9]. It was the second species of pleurocerid snail that Lea ever described,
that stuck [10].
Lea gave the habitat of M. troostiana as “Mossy Creek,
Jefferson County, Ten,” which is quite atypically precise, by 19th
century standards. His description led
with “shell elevated” and followed with “thickly striated.” But in his remarks, he focused primarily on
the “sharp carina” demonstrated by the shell, and secondarily on the “numerous
striae,” which reminded him of what we call today Pleurocera virginica.
If you’re not entirely sure about the distinction between a
carination and a striation, click the image below for a primer on shell
morphology.
I speculate that the nomen “Melania troostiana” receded into
obscurity in the 20th century for two reasons. First, Lea’s 1838 figure doesn’t match any
pleurocerid that currently lives or ever has lived in the state of Tennessee
especially well, particularly with respect to that dramatic carination that
extends from the juvenile into the adult whorls. Hence all subsequent authors have restricted
its range to Mossy Creek.
 |
| Pleurocera troostiana (Click for shell terminology) |
And second, Mossy Creek is a scummy mess. I first had the occasion to visit that
unfortunate little body of water in the summer of 2011, when I was surveying
the Tennessee drainage above Chattanooga for the FWGTN web resource. The Mossy Creek catchment is a mixture of
overly-grazed pastureland and dusty rock quarries, draining directly through
Jefferson City into Cherokee Reservoir.
The creek has obviously suffered decades of erosion, sedimentation, and
enrichment. And at none of the (rather
few) points of access did I find any pleurocerids whatsoever.
My resolve was reinforced, however, by the troostiana figure
in Burch [7], which looked both modern and plausible. The UMMZ does hold three lots identified as
P. troostiana collected from Mossy Creek by somebody named “Andrews.” I don’t see any collection dates in their
online database, but it seems possible to me that Andrews might have been a
Goodrich contemporary, and hence his samples might be relatively recent. And most interestingly, Burch’s figure,
almost certainly from one of those UMMZ lots, does not show that weirdly strong
carination.
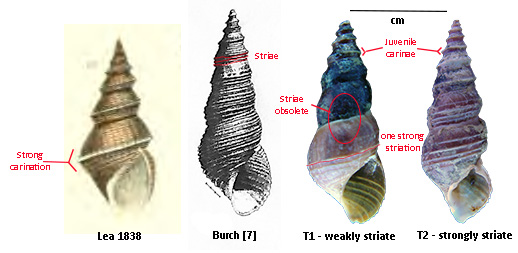
So I returned to Mossy Creek this past spring, and by dint
of persistence was able to find exactly N = 6 topotypic specimens of P.
troostiana at the Old Andrew Johnson Highway bridge (36.1272, -83.4862). The shells borne by all six of those
specimens were striate, although in one case (T1, above) the striae became
obsolete in spots. Three of the shells
matched the figure in Burch quite closely.
And shells of two individuals (T2, above) were so strongly striate that
a carination developed reminiscent of Lea’s original figure. figure.
Seven paragraphs ago I wrote that “the” thing that matters to
us today is that the nomen, “Melania troostiana” was early. Let me modify that slightly. The earliness of the taxonomic act is
certainly important. But just as
important is the tremendous variability in shell striation depicted in
the figure above. All four of those
shells were borne by a single population of snails inhabiting a single little ten
km creek in East Tennessee. Let that
sink in a minute.
Goodrich devoted the entirety of paper number V in his
“Studies of the Gastropod Family Pleuroceridae” series [12] to documenting the
“transient, sporadic” character of shell “spirals,” or striations. Selecting “Goniobasis porrecta Lea of the big
hillside spring at Cumberland Gap” as one of his many examples, Goodrich
observed:
“No multistriate specimens have been seen as from the type locality, but such individuals amount to 32.4 per cent of seventy-seven shells taken from Gap Spring Creek about four miles below the spring.”
Hey kids, test your memory! We devoted our entire essay back in August to
cryptic phenotypic plasticity in the pleurocerid populations of Gap Spring Creek. With what name did I identify those
populations that Isaac Lea described in 1863 as Goniobasis porrecta? Answer at footnote [13] below! Next month, we’ll find out why.
Notes:
[1] The biographical details for this month’s essay, as well
as the striking figure, were extracted from a “Sketch of Gerard Troost,”
published anonymously in the June, 1894 issue of The Popular Science Monthly,
pp 258 – 264.
[2] It was Currey who sent Isaac Lea the sample of
pleurocerids from “Robinson County, Tenn” he described as Melania ebenum in
1843. See:
- CPP Diary: What is Pleurocera (aka Melania, aka Goniobasis, aka Elimia) ebenum? [3Oct19]
[3] Scudder, N. P. (1885)
Bibliographies of American naturalists – II. The published writings of
Isaac Lea, LL.D. Bull. US National
Museum 23: 1 – 278.
[4] Lea, Isaac (1838-39) Description of New Freshwater and
Land Shells. Transactions of the
American Philosophical Society (New Series) 6: 1 – 154.
[5] Tryon, G. W. (1873)
Land and Freshwater shells of North America Part IV, Strepomatidae. Smithsonian Miscellaneous Collections 253: 1
- 435.
[6] Goodrich, C. (1940) The Pleuroceridae of the Ohio River
drainage system. Occas. Pprs. Mus. Zool.
Univ. Mich., 417: 1-21.
[7] This is a difficult work to cite. J. B. Burch's North American Freshwater
Snails was published in three different ways.
It was initially commissioned as an identification manual by the US EPA
and published by the agency in 1982. It
was also serially published in the journal Walkerana (1980, 1982, 1988) and
finally as stand-alone volume in 1989 (Malacological Publications, Hamburg,
MI).
[8] Graf, D. L. (2001)
The cleansing of the Augean stables.
Walkerana 12(27): 1 - 124.
[9] The first valid species of pleurocerid snail described
by Isaac Lea, by the Goodrich/Burch canon, was Melania acuta (Lea 1830). This is not the Pleurocera acuta of Say
(1821). This is a “longitudinally
undulated and transversely lineated” North Alabama species attributed to
Goniobasis by Goodrich, or Elimia by Burch. It may be a synonym of laqueata (Say 1829). Or it is possible that Lea's (1830) acuta is a senior synonym of what we are calling troostiana (Lea 1838), in which case we'd have to salt it and butter it as a double-predestinarian hominy under acuta (Say 1821). Crap, I hate this sort of stuff, and am clean out of patience with it.
[10] Nineteen of the 38 canonical pleurocerid taxa older
than 1838 were described by Lea’s nemesis, Timothy Abbott Conrad, which must
have ticked him off royally. Another 12
were from Thomas Say, whom Lea didn’t much care for either, apparently. See last month's post:
- Isaac Lea drives me nuts [5Nov19]
[11] I’ll post a dot-map showing the site of my Mossy Creek
collection next month.
[12] Goodrich, C. (1935)
Studies of the gastropod family Pleuroceridae – V. Occasional Papers of the Museum of Zoology,
University of Michigan 318: 1 – 12.
[13] I identified the Gap Creek population of G. porrecta as
Pleurocera troostiana, of course! That’s
where I’ve been going with this entire essay.
Shame on you for reading this footnote. Go back and read:
- CPP Diary: Yankees at The Gap [4Aug19]



















