Quite a few malacological mysteries have their origins in the life and work of Thomas Say, the "Father of American Malacology" (1). From his high perch among the founders of the Academy of Natural Sciences in Philadelphia, Say described many of the most widespread and familiar mollusks in the New World. Yet, as we saw in our June '08 essay on Lymnaea humilis (2), his descriptions were spare, his figures few and poor, and his original collections lost. So in this, the second installment of an occasional series, we'll pick up our magnifying glasses and sleuth our way through another dark and tangled maze, tracking the true identity of Say's mysterious Physa heterostropha. Along the way we'll encounter another striking example of the incompatibility between science (the construction of testable hypotheses about the natural world) and law (as represented by the international code of zoological nomenclature.)
The biology will not be at issue here. The best evidence available at present suggests that two morphologically variable species of the genus Physa range across the length and breadth of North America, one bearing a convex shell apex and a two-part penial sheath, the other bearing a concave shell apex and a one-part penial sheath (3). Both have been described and re-described many times, accumulating dozens of aliases in the process.
The concave species was first described by Draparnaud in 1805, not here in America where it is native, but rather as an introduced species in France. The earliest name for this physid, which I have nominated (4) as "the world's most cosmopolitan freshwater gastropod," is Physa acuta. The second-oldest name available for American physids is Thomas Say's (1817) Physa heterostropha (5), which F. C. Baker (6) nominated to the post of "most misunderstood mollusk in America." And the third-oldest name is Say's (1821) Physa gyrina (7), clearly and unambiguously bearing a convex shell apex.
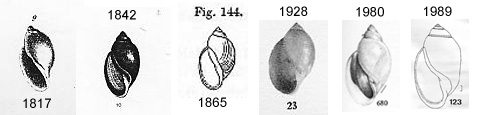 Thomas Say's written description of "Lymnaea" (later Physa) heterostropha (8) stated that the apex is "acute," which might suggest that he was holding a shell of the concave species in his hand on that fateful day in 1817. But his small and poorly-detailed figure (at only 17 mm certainly intended as a 1:1 representation) appears to show a convex apex (at far left above). And the type locality ("Delaware River") might be inhabited by either species.
Thomas Say's written description of "Lymnaea" (later Physa) heterostropha (8) stated that the apex is "acute," which might suggest that he was holding a shell of the concave species in his hand on that fateful day in 1817. But his small and poorly-detailed figure (at only 17 mm certainly intended as a 1:1 representation) appears to show a convex apex (at far left above). And the type locality ("Delaware River") might be inhabited by either species.If the nomen "heterostropha" is correctly applied to the species of American physid with a concave apex, it is a junior synonym of acuta, and disappears. But if heterostropha is applied to the convex physids, it would be the senior synonym of gyrina, and by priority the correct name for a species of freshwater snail widespread across North America.
Modern scholarship has cast doubt on whether Say's type specimens actually remain in the collection of the Academy of Natural Science in Philadelphia. But there exist today not one but two very old lots of Physa heterostropha in the ANSP collections which for many years were believed to have originated with Thomas Say (9). They bear no data of any sort beyond their Latin binomena, and they contain specimens bearing both concave and convex apexes.
The first monographic review of the American Physidae was that of S. S. Haldeman (1842), Say's successor at the ANSP (10). Haldeman figured 19 Physa heterostropha shells on two plates, including (to my eye) six with convex apexes and 13 with concave. The next monograph was that of Binney (1865) who (apparently quite randomly) selected one shell with a very concave apex and figured it over the label, "Physa heterostropha, from Say's type" (11).
Frank Collins Baker was the first to document the striking difference in the penial morphology of the concave and the convex physids, working in Wisconsin in 1928 (6). He gathered 5 species of the former group into the subgenus Physodon, bearing a one-part penial sheath, leaving 15 convex species in the subgenus Physella (s.s.), with penial sheaths divided into two parts. Baker tentatively listed heterostropha among the Physella (s.s.), on the basis of the convex shells of the specimens he had personally collected in the Philadelphia area. He did not consider that the species ranged into Wisconsin, however, and hence admitted no direct observations of its penial morphology.
Our modern concepts of the physid taxa are largely due to the work of George Te (12), as reproduced in Burch's (1980) "North American Freshwater Snails" (13). Although the origin of Te's sample of P. heterostropha is not clear, his concept of the species as bearing a concave apex and a one-part penial sheath has come to predominate in the years that have followed. This is the understanding of P. heterostropha that my colleagues and I brought to our research synonymizing heterostropha under P. acuta (4).
But there is yet one additional coil in the serpentine history of America's most misunderstood mollusk. For astute readers will note that at no time during its first 172 years in the scientific literature did anybody formally designate a lectotype for Say's Physa heterostropha.
That solemn duty was assumed by Shi-Kuei Wu in his (1989) inventory of the freshwater mollusks of Colorado (14). Wu wrote, "The two type lots housed at the Academy of Natural Sciences of Philadelphia was actually (sic) mixed lot (together with Physa gyrina) and had not been positively identifiable (Baker, 1964). After examining these two type lots, I have concluded that this confusion can best be rectified if the specimen figured by Haldeman (1842) on Plate 1, fig. 10 in his monograph should be designated as lectotype. That specimen (ANSP 280031) is hereby so designated."
It is not clear why Wu picked the tenth of Haldeman's 19 figures of P. heterostropha, nor is the match between Wu's lectotype and Haldeman's #10 especially convincing, as can be seen in the group figure above (labeled 1842 and 1989). It is quite clear, however, that Wu selected an extremely convex specimen, effectively synonymizing gyrina under Physa heterostropha. And in identifying the other specimens in the mixed lots - those with concave apexes - as "Physa gyrina," he introduced a new confusion that I don't think existed prior to 1989. He got the backwards snails completely backwards.
Wu went on to recognize eight species of Physa in Colorado, about half of which he reversed. He assigned the names gyrina and anatina to concave shells, while in the larger community those names are typically associated with convex populations, and assigned the names heterostropha and integra to convex shells, when most workers would associate those names with concave populations. This confusion carried into the 1997 "Missouri Aquatic Snails" book of Wu, Oesch and Gordon (15), which included 12 species of Physa, gyrina re-interpreted to its original position with the convex group but heterostropha and anatina still backward. And at least one allozyme paper was also published with the backwards snails backwards, that of Liu in 1993 (16).
I am not an attorney, but my reading of the International Code of Zoological Nomenclature suggests to me that Wu's (1989) type designation may be valid. His concept of heterostropha is defendable all the way back to 1817, and given its not-infrequent use in the subsequent literature, would seem unlikely to be overturned on appeal. But as a scientist, it seems clear to me that the name "heterostropha" has become worse than useless - it is an actual impediment to our understanding of the evolutionary history of an important group of organisms. To substitute an ambiguous name like "heterostropha" for the much cleaner name "gyrina" makes no practical sense.
So the bottom line is that I intend to call the concave species acuta, the convex species gyrina, and consign heterostropha to the dustbin. Add another line to my lengthy rap sheet in the file cabinet at the ICZN squad room. And when the taxonomy police come to surround the building with guns and dogs, well, let's just say ... They'll never take me alive!
P.S. From Gary Rosenberg
Date: Tue, 28 Oct 2008 13:32:25 -0400
To the FWGNA group:
Those of you with an unquenchable thirst for malacological mystery may enjoy my email exchange with Gary Roseberg below, regarding the type of Physa heterostropha. In our latest twist, The Butler (yours truly) has been cleared of all charges. But the mystery deepens. Did Thomas Say's holotype ever reside in the ANSP? If not, what deceived such clever investigators as W. G. Binney, H. A. Pilsbry, and H. B. Baker? And if so, what has become of that type material today? Has there been foul play?
Only The Shadow knows
Rob
Subject: Re: Backwards Snails Backwards!
Date: Tue, 14 Oct 2008 14:34:22 -0400
From: "Gary Rosenberg"
To: "Dillon Jr., Robert T"
ICZN Police! Come out with your head up!
I'm please to inform you, Dr. Dillon, that the charges against you have been dismissed. The lectotype designation by Wu has no standing, because there is no evidence that the Haldemann material was studied by Say. ANSP 280031 was donated by Haldemann, and is labeled as Haldemann's types. This means that they are Haldemann's figured specimens, not types of earlier named species.
Best wishes,
Gary
Subject: RE: Backwards Snails Backwards!
Date: Tue, 14 Oct 2008 14:47:08 -0400
From: "Dillon Jr., Robert T"
To: "Gary Rosenberg"
Dear Gary,
Good to hear from you, old buddy.
So do you have any idea why everybody for 150 years considered that particular lot or lots to have been Thomas Say's, and conversely, how we have now decided that it isn't? And when did this reversal of opinion occur? Is this your own insight, or that of some other scholar, and on what evidence was this conclusion based? Has some sort of "retraction"of H. B. Baker been published, or is this just an informal understanding?
Thanks for the reprieve,
Rob
Subject: RE: Backwards Snails Backwards!
Date: Tue, 14 Oct 2008 18:12:59 -0400
From: "Gary Rosenberg"
To: "Dillon Jr., Robert T"
Hi Rob,
>>>So do you have any idea why everybody for 150 years considered that particular lot or lots to have been Thomas Say's... We don't even know that; we don't know what lots people were referring to, as explained further below. >>> and conversely, how we have now decided that it isn't? And when did this reversal of opinion occur? Is this your own insight, or that of some other scholar, and on what evidence was this conclusion based? Has some sort of "retraction" of H. B. Baker been published, or is this just an informal understanding?
This is my own interpretation. Unfortunately, Baker didn't state the catalog numbers of the lots he thought came from Say. Haldemann's figured specimens were not catalogued until 1962 (hence numbers in the 280,000s). I don't know if they were in the collection before then, but other Physas of that vintage was catalogued around 1915. (We didn't start assigning catalogue numbers here until the 1890s.) It is strange that the Haldemann material was catalogued so late. It could be that it resurfaced later, or it might have been left in the collection uncatalogued because it contained multiple species. Then, it preparation for Baker's type catalogues, numbers were assigned later.
The Haldemann material was originally glued onto boards, with specimens arranged as figured on the plates. Most of the specimens have since been removed from the boards (which are kept with ANSP 280031). There are two lots from Haldemann containing material he identified as P. heterostropha, one for plate 1, the other for plate 2. It is possible that Baker (1964) meant these lots when he referred to our having two lots from Say. But our catalog does not state that these are type lots and the labels do not state that they are types of particular species. Yet specimens from the same set of boards were catalogued at the same time and were entered as types in our catalogue.
Haldemann did not state that he had examined material of P. heterostropha from Say, but he did note Say connections for several other species. For example, Haldemann obtained specimens of P. gyrina and L. caperata from Mrs. Say and he examined L. obrussa, Amnicola lustrica and Paludina transversa in the Academy's collection. If Say's material was at the Academy, it would have been unusual for Haldemann to combine it with his own.
So I conclude:
a) There is no evidence that Haldemann had access to Say's material of P. heterostropha. Wu's (1989) lectotype designation is therefore invalid.
b) The only evidence that Haldemann's lots of P. heterostropha are the two lots that Baker referred to is that they are no other Say lots that he might have been referring to. So either there are two lots now missing from the ANSP collection, or Baker was referring to the Haldemann material.
c) Binney (1865) refers to types of P. heterostropha at ANSP and illustrates one. Pilsbry (1894, "Critical list of mollusks collected in the Potomac Valley. PANSP 46: 11-31) refers to "the type". These specimen(s) can no longer be identified. If they correspond to the Haldemann material they are not types; if they are correspond to other lots, they have been lost. Either way, no type material exists for P. heterostropha.
d) If Says' description is not sufficient to identify the species, then a neotype is needed. By the way, the type locality is not the Delaware; Say (1817) said "Inhabits the Delaware river and many other waters of the United States....", so a neotype could come from anywhere in the US.
Maybe this should be written up formally?
Best wishes,
Gary
Notes
(1) The Wikipedia entry on Say would benefit from a contribution by the malacological community.
(2) Malacological Mysteries I: The type locality of Lymnaea humilis. [25Jun08]
(3) Wethington, A.R. & C. Lydeard (2007) A molecular phylogeny of Physidae (Gastropoda: Basommatophora) based on mitochondrial DNA sequences. Journal of Molluscan Studies 73: 241 - 257. A PDF download is available from our October '07 on the classification of the Physidae.
(4) Dillon, R. T., A. R. Wethington, J. M. Rhett and T. P. Smith. (2002) Populations of the European freshwater pulmonate Physa acuta are not reproductively isolated from American Physa heterostropha or Physa integra. Invertebrate Biology 121: 226-234. [PDF]
(5) Say, T. (1817) The article, "Conchology" from the 4th volume of the American edition of Nicholson's British Encyclopedia of Arts and Sciences, Philadelphia.
(6) Baker, F. C. (1928) Freshwater Mollusca of Wisconsin, Part I, Gastropoda. Bull. Wisc. Geol. Natur. Hist. Survey, no. 70. Madison: University of Wisconsin Press. I published an appreciation of F. C. Baker in November '06.
(7) Say, T. (1821) Descriptions of Univalve Shells of the United States. Journal of the ANSP 2:172.
(8) "Lymnaea heterostropha - Shell sinistral, subovated; color, pale yellow, chestnut or blackish; whorls four, the first large, the others very small, terminating rather abruptly in an acute apex; aperture large, somewhat oval, three-fourths of the length of the shell, or rather more; within of a pearly lustre, often blackish; lip a little thickened on the inside, and tinged with dull red." (Say, 1817)
(9) Baker, H. B. (1964) Type land snails in the Acady of Natural Sciences of Philadelphia, Part III. Limnophile and thalassophile Pulmonata. Proc. Acad. Nat. Sci. Philadelpha 116: 149 - 193.
(10) Haldeman, S. S. (1842) A monograph of the Freshwater Univalve Mollusca of the United States, Physadae. E. G. Dorsey, Philadelphia. 40 pp.
(11) Binney, W. G. (1865) Land and Fresh Water Shells of North America, Part II. Pulmonata Limnophila and Thalassophila. Smithsonian Misc. Collections No. 143. 161 pp.
(12) Te, G. A. (1978) The systematics of the family Physidae (Basommatophora: Pulmonata). Ph. D. Dissertation, University of Michigan. 325 pp
(13) Burch, J. B. (1980, 1982, 1989) North American Freshwater Snails. Malacological Publications, Hamburg, MI.
(14) Wu, Shi-Kuei (1989) Colorado Freshwater Mollusks. Natural History Inventory of Colorado, Number 11. 117 pp.
(15) Wu, Shi-Kuei, R. D. Oesch & M. E. Gordon (1997) Missouri Aquatic Snails. Missouri Department of Conservation, Natural History Series Number 5. 97y pp.
(16) Liu, H-P. (1993) Diagnostic genetic loci for species in the genus Physella. Malac. Rev. 26: 1 - 8.
 Although the natural habitat of the NZMS is generally considered to be the muddy margins of lakes and rivers, in the media it is almost always associated with pocket change.
Although the natural habitat of the NZMS is generally considered to be the muddy margins of lakes and rivers, in the media it is almost always associated with pocket change.  Media attention seems to have played an important role in mobilizing public sentiment. In late June, an invasion by "harmful snails" or "worrisome snails" was the subject of several television news stories and reports in the
Media attention seems to have played an important role in mobilizing public sentiment. In late June, an invasion by "harmful snails" or "worrisome snails" was the subject of several television news stories and reports in the 

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}