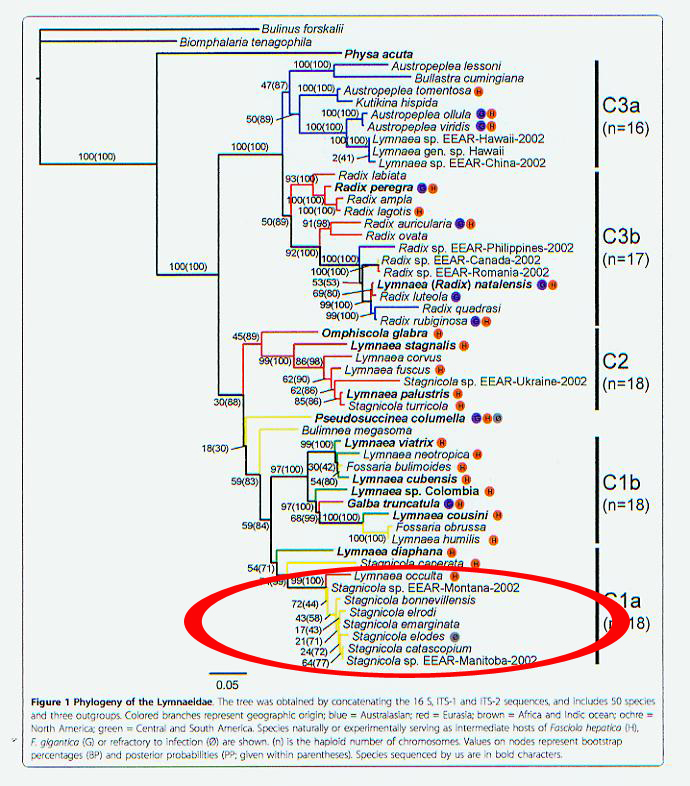
In June we reviewed the 2010 paper by Ana Correa and her colleagues synthesizing 15 years of effort (by several independent research groups) to develop gene trees for the Family Lymnaeidae [1]. Reasoning from known levels of interpopulation divergence in Lymnaea stagnalis, it appeared to us that approximately 5% sequence divergence may be expected among conspecific lymnaeid populations sharing the same continent, and 10% between continents. By that "stagnalis yardstick," the gene tree of Correa seemed to evince approximately 19 species worldwide. That's approximately what Bengt Hubendick would have suggested in 1951 [2].
Correa and her colleagues were not, however, primarily motivated by an academic interest in the evolution of the Lymnaeidae. Their explicit motive was to develop a "phylogenetic framework" upon which to "analyze how susceptibility to infection by Fasciola hepatica and F. gigantica has evolved [3]." Liver flukes of the genus Fasciola cause significant losses of livestock worldwide, and (especially in Latin America) may be sporadically recorded in human populations as well.

And so we should not be surprised that Correa and colleagues followed their 2010 study with a second study published in 2011 [4], focusing on the neotropical lymnaeid fauna, almost all of which are what we here in temperate North America traditionally call "fossarines."
The fossarines are small-bodied lymnaeids typically found living amphibiously at the edges of ditches, ponds, and quiet backwaters. They are common inhabitants of wet pastures and the trampled margins of riverbanks where livestock come to water, and as such have become the most important hosts of fascioliasis. Click the figure above and spot the deer hoof print [5].
European populations of this type are usually identified simply as "Galba" truncatula. But here in North America their taxonomy is much messier. The Burch/Baker system [6] lists approximately 12 species of "Fossaria" in two subgenera, the bicuspid subgenus "Bakerilymnaea" with cubensis, bulimoides, and several others, and the tricuspid subgenus "Fossaria s.s." with humilis, modicella, parva, obrussa, and several others, including truncatula from Europe. Hubendick lumped all our North American tricuspid nomina under humilis, keeping two names for our bicuspid populations: cubensis in the east and bulimoides in the west. He also admitted truncatula to the North American fauna, extending only through Alaska and a bit of Canada.
In South America Hubendick recognized viator (which he found difficult to distinguish from cubensis), cousini, and pictonica, a nomen that seems to have become lost. But in contrast to the situation in temperate North America, the taxonomy of the South and Central American Lymnaeidae has not remained on ice for 30 years. Rather, the availability of funding for fascioliasis research in the tropics has led to the description of a welter of new species, including neotropica, shirazensis, and meridensis, which have been added to the older nomina cubensis, viator/viatrix, cousini, and truncatula as well.
In recent years a malacological football match seems to have developed between French and Spanish teams working on the systematics of these little critters. And in the spirit of full disclosure, I should confess that I have not spent the entire game on the sidelines. In 2008 I was contacted by Prof. Dr. S. Mas-Coma (Universidad de Valencia) to furnish topotypic L. humilis for sequencing work, which led to my post of 25June08 [7] informally "restricting" the type locality of L. humilis to Owego, New York. Ultimately I sent Dr. Mas-Coma samples of both Charleston cubensis and L. humilis from Owego. And in 2009 I sent samples of Charleston cubensis to Dr. Philippe Jarne (CNRS Montpellier) as well [8].
So just as we asked in June for Correa's 2010 paper, we might reasonably ask today for Correa's paper of 2011. Do we know enough about the biology of the little fossarine lymnaeids to make any sense of all the sequence data that have accumulated in recent years? The quick answer is, no. If you've stumbled onto this essay looking for any actual enlightenment regarding the systematics or evolution of the Lymnaeidae, quit now.
Those of you still with me may recall that my June application of our "stagnalis yardstick" to the fossarine branch of Correa's 2010 gene tree returned two groups, which I labeled with three Hubendick-era names, the bicuspid "cubensis" and the tricuspid "humilis/truncatula." Does that fossarine branch comprise three genuinely distinct taxa, two, or one? Let's fish topotypic CO1 sequences out of GenBank for these three old, well-established specific nomina, BLAST them against the entire NCBI nucleotide database, apply our stagnalis yardstick, and see. And we might as well see where all those other (primarily neotropical) taxa fall as well, while we're at it.
For Lymnaea cubensis I used AM49009, collected from Cuba and sequenced by Bargues et al [9]. For L. humilis I used FN182197, collected by myself from the (newly restricted) type locality at Owego, New York, and also sequenced in the Bargues / Mas-Coma lab [10]. For L. truncatula I used EU818799, collected from Germany [11] and sequenced by Albrecht and colleagues [12]. I then ran simple BLAST searches from each of those three topotypic sequences and noted percent sequence divergence across the entire set of matches returned as "significant" by the search tool. The results are shown graphically in the figure below.

Using truncatula as a query sequence, the BLAST search returned 14 sequences in the 96-100% similarity range, all labeled by their collectors as truncatula. In the 90-93% similarity range there were 26 sequences, including one truncatula, all five cousini, all four humilis, all three shirazensis, six (of eight) viator/viatrix, three (of the seven) neotropica, and four unidentified.
Querying from humilis, the BLAST tool returned the three other humilis CO1 sequences in GenBank matching in the 99-100% range. Matching in the 90-94% range were all 16 truncatula, all seven neotropica, all five cousini, all three shirazensis, two (of the five) cubensis, and the four unidentified.
Looking down from cubensis, BLAST returned 15 sequences in the 95-100% similarity range: the other four cubensis, six (of the seven) neotropica, four (of the eight) viator/viatrix, and the single bulimoides sequence [15] in the database. In the 90-94% range were just eight sequences: the other four viator/viatrix, two (of 16) truncatula, one (of five) cousini, and one (of four) humilis.
So the bottom line is that fossarine populations called truncatula in Europe and humilis in North America demonstrate the same level of sequence divergence we have observed between populations of L. stagnalis universally considered conspecific on both continents.
Then if one considers those 16 + 4 sequences together, there is some evidence suggesting that humilis/truncatula may be conspecific with the cubensis-type nomina shown red in the figure above, if one thinks of humilis/truncatula and cubensis as inhabiting separate continents. Some of the 21 cubensis-type sequences demonstrate divergence in the 5-10% range from the 20 humilis/truncatula sequences, others are more divergent. But if one thinks of both humilis and cubensis as sharing a continent, which I suppose I do, then one probably ought to interpret the relationships depicted above as supporting the two-species model, weakly.
In any case, the present analyis contains very little evidence supporting any of the other nominal species shown in the figure above, expecially bulimoides [15], neotropica, and viator/viatrix, all of which demonstrate less than 5% sequence divergence from cubensis. This introduces the further complication that D’Orbigney’s 1835 nomen viator is an older name than Pfeiffer’s 1839 cubensis. I would change the name at the top of the FWGNA page from cubensis to viator if I believed in DNA sequence data, which I do not.
Correa and colleagues [4] go beyond believing in sequence data to a belief in gene trees, a strict and demanding faith indeed. While concurring with us that cubensis, neotropica, and viator/viatrix appear conspecific, their 2011 analysis suggested to them as many as five “species” of fossarines: truncatula, humilis, cubensis, cousini, and (surprise!) a new species from Columbia and Venezuela, which (I feel sure) will reach description soon. They concluded “that conchological and anatomical characters are uninformative [13] to identify closely related species of Lymnaeidae, and that DNA-based approaches should be preferred.”
Yes, trying to advance the ball downfield entirely by head-butting will rarely score points, and so the foot should be preferred. Or, how about using our hands?
As far as I can determine, nobody has ever tried to rear any New World population of fossarine snails in culture, much less gather the first datum on their mating system. Several excellent studies of European truncatula have returned evidence of preferential self-fertilization [14], but I am not aware of any study involving humilis, cubensis, or the scores of genetically similar nominal species of the Americas, temperate or tropical. So big international research teams continue to fill the journals with forests of gene trees, while simple questions about the basic biology of the snails themselves go unaddressed. Such is the sad state of The Lymnaeidae, 2012.
Notes
[3] Come on, guys! In the entire sample of 51 taxa analyzed by Correa et al (2010), only two have ever been demonstrated refractory to infection by Fasciola. Susceptibility is generally distributed all over the entire lymnaeid phylogeny. Look at the little colored balls here: [Correa 2010 tree]
I hate to be cynical, but it's hard to see how research in this direction is going to solve the global fascioliasis problem.
[4] Correa, A.C., J.S. Escobar, O. Noya, L.E. Velasquez, C. Gonzalez-Ramirez, S. Hurtrez-Bousses & J-P. Pointier (2011) Morphological and molecular characterization of Neotropic Lymnaeidae (Gastropoda: Lymnaeoidea), vectors of fasciolosis. Infection, Genetics and Evolution 11: 1978-1988. [Open Access]
[5] Photo taken in September of 2008 at a muddy pool in Owego, NY, the type locality of Lymnaea humilis.
[6] The classification of the Lymnaeidae adopted by Burch for his (1989) "North American Freshwater Snails" was a modification of the system developed by Baker (1911, 1928). For more background, see my post of [28Dec06].
[8] This transaction may have led, in some complicated way, to an error in the 2010 paper by Correa et al. In her Table 1, Correa gave the collection locality for Lymnaea humilis as "USA, Charleston (South Carolina)." No, the three GenBank accession numbers she cited all corresponded to the sequences Bargues & Mas-Coma obtained from my samples of Owego, New York [10]. There are no L. humilis populations in Charleston! That was the entire point of my 25June08 essay.
I understand that the North American situation regarding cubensis and humilis is confusing. But it troubled me to discover that the single fact about which I had any independent information in the Correa 2010 paper was completely bass-ackward wrong.
[9] Bargues, M.D., P. Artigas, R.L. Mera y Sierra, J.P. Pointier & S. Mas-Coma (2007) Characterisation of Lymnaea cubensis, L. viatrix and L. neotropica n. sp., the main vectors of Fasciola hepatica in Latin America, by analysis of their ribosomal and mitochondrial DNA. Ann. Trop. Med. Parasitol. 101: 621-641.
[10] Bargues, M.D., P. Artigas, R.T. Dillon & S. Mas-Coma (unpubl.) Molecular characterization of Lymnaea humilis (= L. modicella), a major fascioliasis vector in North America, and evaluation of the usefulness of nuclear rDNA and mtDNA markers for Lymnaeidae.
[11] Although there were quite a few truncatula sequences in GenBank, most authors gave no data on collection locality, which as I noted in June, drives me nuts. Albrecht and colleagues [12], thank heaven, reported their collection site as "Germany: Thuringia, Erfurt-Bindersleban, spring of Nesse River." I have no idea how close this might be to the actual type locality, given by Muller as "Thangelstedt, near Weimar, Germany," but we'll make it do.
[12] Albrecht, C., C. Wolff, P. Gloer & T. Wilke (2008) Concurrent evolution of ancient sister lakes and sister species: the freshwater gastropod genus Radix in lakes Ohrid and Prespa. Hydrobiologia 615: 157-167.
[13] The Correa et al. 2011 paper also included a nice dataset on penial morphology, but really very little on the shell - just simple length and width. And none of the European workers ever seem to examine radular morphology. I have no idea why not.
[15] Note added 13Feb24. The "single bulimoides sequence" referred to above is spurious, and Lymnaea bulimoides is not a junior synonym of L. cubensis/viator. For more, see:
- What is Lymnaea bulimoides [13Feb24]