It's not unusual for retail stores to open their doors for business, operate for a few weeks, and then announce a "Grand Opening." I'm not sure about the rationale for this practice, but welcome to the Grand Opening of the FWGNA Blog! Doing business since July, 1998.
For any newcomers who might stumble onto this post - the Freshwater Gastropods of North America project was born at the first World Congress of Malacology in Washington, 12 years ago this summer. Our objective has always been to survey the entire continent for freshwater snails, by any means necessary. Four states mostly done as of February 2010! What a team!
In any case, very early in the effort I fell into the habit of sending mass emails to an ever-growing addressbook of colleagues - 186 at last count - recently about once per month*. Often these messages have been simple news items about upcoming meetings, grants, employment opportunities, or whatever. But increasingly I've found myself reviewing and reporting on the current literature, offering opinions on taxonomic questions, and pontificating on invasive species, endangered species, and matters at the interface between science and public policy.
Clearly we've needed a medium to open up a better conversation for a long time. I'm not sure why I didn't migrate to the blog format years ago.
An important (although perhaps secondary) benefit of modern blogging is the convenience of the indexing function. So over the last couple weeks I've uploaded (from the "FWGNA Archives") the great majority of my old email posts, "tagging" them as I did with the brief descriptors you see listed at right. And I imagine you've noticed the Google-powered search box. It may take a while for the Google webcrawlers to find this tiny corner of the internet and "populate" that search box, but eventually it should become a helpful tool.
And now its up to all of us to give those web-crawlers something to feed on! Regarding comments - I am advised that, unless I moderate this blog, unscrupulous agents will use the blank spaces provided to advertize viagra. But you do not need a Google account to comment. Type your message in the comment box, and choose "name" in the "select profile" box, or even "anonymous" if you want. I'll get an email alerting me that a comment needs my approval, and do so promptly.
Looking forward to it,
Rob
*I'm still planning to maintain my email list, and I guess I'll continue to send monthly emails. But they'll be short - probably just a title and a link to the blog.
Sunday, February 28, 2010
Wednesday, January 20, 2010
Okay, okay! I'll start a blog!
To the FWGNA group:
My message of 18Dec09 regarding the community consequences of Bellamya invasion seems to have struck a chord. In addition to the scattering of "atta-boys" I often receive (and always appreciate), last month's post attracted four comments that seem substantive enough to share.
John Havel (of Missouri State) wrote me with several insights regarding invasion biology in lakes (as applied to Bellamya in particular), David Campbell (Alabama) offered taxonomic comments, and both Jeremy Tiemann (Illinois Natural History Survey) and Bob Prezant (Montclair) suggested additional references. I have appended (slightly edited) versions of the emails I received from these four gentlemen to the end of my 18Dec09 post.
When I first started sending messages to the FWGNA group back in July of 1998, I had never heard the word, "blog." But I just googled-up Wikipedia, and apparently the term itself dates to 1994. The first "hosted blog tools" became available in 1999.
In recent years, the internet has become something more like a community, for better or worse. I have always, at least occasionally, posted your comments regarding my monthly essays in the form of a "P.S." in the FWGNA archives. But this is an old-timey pairwise approach, where I say something to you, and then you say something to me. The internet now seems to have developed to a point where everybody expects to talk to everybody else, as a community.
OK, okay - I'll start a blog. I don't know how I'll go about doing this, or when the transformation will occur. But the advantages of group communication are as obvious to me as they must be to everybody I'm talking to right now, pairwise.
So we'll keep in (better) touch,
Rob
Friday, December 18, 2009
Community Consequences of Bellamya Invasion
Editor’s Note – This essay was subsequently published as: Dillon, R.T., Jr. (2019d) Community consequences of Bellamya invasion. Pp 47 - 53 in The Freshwater Gastropods of North America Volume 4, Essays on Ecology and Biogeography. FWGNA Press, Charleston.
I dearly love experiments. Does it seem to anybody else on this list that, over the last 20-30 years, the experiment is going out of style? In any case, the recently published research results of Pieter Johnson, Chris Solomon, and their colleagues on the community effects of Bellamya (1) invasion, which include both experimental and field components, arrive as especially welcome contributions to our (surprisingly slender) file on the biology of invasive viviparids.
Bellamya populations (2) seem to have appeared in the Northern Highlands lake district of Wisconsin sometime in the 1950s. Johnson and his colleagues (3) conducted an 8-week experiment in a set of 24 outdoor “mesocosms” containing 1,000 of water from nearby Sparkling Lake, a clean sand/gravel substrate (inoculated with lake mud), two cinder block “refugia,” and 45 individuals each of Physa gyrina, Lymnaea stagnalis, and Helisoma trivolvis. Eight of the mesocosms received 45 Bellamya, eight received a pair of crayfish (4), eight received both Bellamya and crayfish, and eight were held as controls.
 The most striking result of these experiments was that the populations of Physa and Lymnaea grew in the absence of Bellamya, but declined in the presence. The control Physa roughly doubled in population size over the 8 weeks, while the control Lymnaea roughly doubled in wet mass, although apparently did not reproduce (5). But all pulmonate populations declined by all measures in the Bellamya treatments. Clearly the invasion of Bellamya through the lakes of Northern Wisconsin should have negative effects on the native freshwater gastropods, right?
The most striking result of these experiments was that the populations of Physa and Lymnaea grew in the absence of Bellamya, but declined in the presence. The control Physa roughly doubled in population size over the 8 weeks, while the control Lymnaea roughly doubled in wet mass, although apparently did not reproduce (5). But all pulmonate populations declined by all measures in the Bellamya treatments. Clearly the invasion of Bellamya through the lakes of Northern Wisconsin should have negative effects on the native freshwater gastropods, right?
Well, of course it’s more complicated than that. The Helisoma populations declined in both biomass and abundance (5) in all mesocosms, including the controls. Whether the Helisoma were outcompeted by the Physa and Lymnaea, or whether these mesocosms were simply unsuitable habitat for Helisoma, we'll never know. My intuition suggests to me that if the researchers had floated some macrophytic vegetation on the surface of their mesocosms, their Helisoma populations would have been fine (6). The problem is that the experiments of Johnson and colleagues show Helisoma going to extinction in the absence of Bellamya just as clearly as they show Physa and Lymnaea going to extinction in the presence (7). As much as I love experiments, a 5’ x 5’ structural foam plastic tank and a lake are two entirely different things.
Who could disagree? Certainly not our colleagues in Wisconsin. Thus in a companion study, Chris Solomon and a gang of four (including yours truly) surveyed 42 lakes in northern Wisconsin, roughly half of which had been invaded by Bellamya and half not, collecting a great variety of environmental measures in the process (8).
We found evidence of positive correlations between the presence of Bellamya and several general measures of lake productivity, such as conductivity and Secchi turbidity, but very little evidence of an effect of Bellamya on the native freshwater gastropod fauna. At neither the site level nor at the lake level could we find any evidence that Bellamya presence or abundance had any effect on the native freshwater gastropod community at all.
The absence of any significant results whatsoever neatly confirms my hypothesis, of course. Casting an eye back through the FWGNA archives, it looks as though I’ve authored fully 13 previous posts on invasive species, primarily Pomacea, Potamopyrgus, and Bellamya, with other viviparids and Bithynia making cameo appearances. And it may be recalled that I am an advocate of the “empty niche” hypothesis, which dates me back to the 1970s with most embarrassing accuracy.
Essentially, I think that the two best predictors for the success of a potential invasion are that the invading population must be weedy and different (9). Invaders must be adapted to the new environment they are invading, of course, while ecologically different from the native inhabitants – the more different, the better.
The gigantic, filter-feeding, ovoviviparous Bellamya is indeed strikingly different from all the other elements of the freshwater gastropod community native to northern Wisconsin. Hence I would not expect to uncover any competitive effects, and feel quite validated that we did not find any.
But I hasten to add two asterisks. First, populations of four different species of viviparids inhabit the lakes of northern Wisconsin, three of which are gigantic, filter-feeding, ovoviviparous invaders: Bellamya chinensis, B. japonica, and Viviparus georgianus. That our distributional data returned no evidence of a negative relationship within this subset, over 197 sites in 42 lakes, does indeed seem to suggest that our methods may have been too weak to detect even bona fide competition, where it occurs.
Second, a special challenge attends the selection of any invasive species for a study of population biology. What would make the researcher think that any such population has reached the carrying capacity of its environment? Competition cannot begin until some resource becomes limiting. If both Bellamya and Viviparus populations are still growing and spreading through northern Wisconsin in 2009, there is no reason to imagine that either will have any effect on the other.
And in fact, our analysis did return evidence of correlations between Bellamya presence and such measures of disturbance such as boat landings and shoreline housing. This implies to us that Bellamya populations may indeed be actively growing and spreading even in northern Wisconsin, where the invasion may have been ongoing for as much as 50 years.
I conclude with a call for additional research (10). This is a button you have rarely seen me push in this series of essays, both because it is trivially obvious to me that we need additional research on all aspects of freshwater gastropod biology, and because I try not to preach to the choir.
But where other people see "invasive species," I see "model organism." We have been greatly enriched, for example, by a huge volume of first-rate research on Pomacea in recent years (11), and that literature is huge (12). Meanwhile I haven't seen more than a couple papers published on even the most basic aspects of the life history of Bellamya, in total, over the 100-year history of the North American invasion (13). And three paragraphs ago I found myself offering an hypothesis regarding carrying capacities of invasive viviparids without a single estimate of population size, even static, ever taken anytime, anywhere. As easy as invasive viviparids are to sample, sitting on clean sand at the bottom of bathtub reservoirs? Shame on us all!
Keep in touch, everybody!
Rob
Notes
(1) I follow Smith (2000) in preferring the (internationally-recognized) generic nomen "Bellamya" over the (provincial) "Cipangopaludina." See the FWGNA page on B. japonica for details and references.
(2) Wisconsin populations are almost entirely Bellamya chinensis. I did identify B. japonica in five (of 42) lakes surveyed, which were lumped with B. chinensis for analysis. That's B. chinensis on the left, and B. japonica on the right.

(3) Johnson, P. J., J. D. Olden, C. T. Solomon, and M. J. Vander Zanden (2009) Interactions among invaders: community and ecosystem effects of multiple invasive species in an experimental aquatic system. Oecologia 159: 161-170.
(4) The crayfish were the invasive Orconectes rusticus. Those results were interesting as well, but don’t bear directly on the subject of this essay.
(5) Pulmonate juveniles are born at approximately 1 mm shell length. The sampling problems of finding such “speck babies” in a 1,000 liter mesocosm with sand and cinder blocks should be obvious to everybody. Thus I strongly suspect the “pulmonate abundance” figures reported by Johnson and colleagues are systematic underestimates.
(6) Helisoma is certainly quite commonly observed on floating vegetation. Any sort of additional habitat along these lines would certainly have benefited all three pulmonate populations.
(7) And if there’s a crayfish in the lake, all pulmonate populations are immediately doomed.
(8) Solomon, C. T, J. D. Olden, P. T. J. Johnson, R. T. Dillon and M J. Vander Zanden (2010) Distribution and community-level effects of the Chinese mystery snail (Bellamya chinensis) in northern Wisconsin lakes. Biological Invasions 12: 1591 - 1605. [PDF] 476kb.
(9) "Invaders Great and Small" Post of September '08.
(10) This should not be construed as a call to sequence two genes from single individuals sampled from 40 imaginary viviparid species in a vain attempt to reconstruct evolutionary events of 100 million years ago, while remaining arrogantly clueless regarding the basic population genetics of even one crappy pond full of crappy snails today.
(11) "Ampullariids star at Asilomar" Post of August '05.
(12) "Review: Global Advances in Golden Apple Snails" Post of May '07.
(13) Stanczykowska, A, E. Magnin and A. Dumouchel (1971) Etude de trois populations de Viviparus malleatus (Reeve) de la region de Montreal. I. Croissance, fecondite, biomasse et production annuelle. Can. J. Zool. 49: 1431-1441. Jokinen, E.H. (1982) Cipangopaludina chinensis (Gastropoda: Viviparidae) in North America, review and update. Nautilus 96: 89-95. Therriault, T. W. and E. Kott (2002-3) Cipangopaludina chinensis malleata (Gastropoda: Viviparidae) in southern Ontario: An update of the distribution and some aspects of life history. Malac. Rev. 35-36: 111-121.
Comments
(1) From John Havel JohnHavel@MissouriState.edu
(2) From David Campbell amblema@bama.ua.edu
(3) From Jeremy Tiemann jtiemann@inhs.uiuc.edu
(4) From Bob Prezant prezantr@mail.montclair.edu
I dearly love experiments. Does it seem to anybody else on this list that, over the last 20-30 years, the experiment is going out of style? In any case, the recently published research results of Pieter Johnson, Chris Solomon, and their colleagues on the community effects of Bellamya (1) invasion, which include both experimental and field components, arrive as especially welcome contributions to our (surprisingly slender) file on the biology of invasive viviparids.
Bellamya populations (2) seem to have appeared in the Northern Highlands lake district of Wisconsin sometime in the 1950s. Johnson and his colleagues (3) conducted an 8-week experiment in a set of 24 outdoor “mesocosms” containing 1,000 of water from nearby Sparkling Lake, a clean sand/gravel substrate (inoculated with lake mud), two cinder block “refugia,” and 45 individuals each of Physa gyrina, Lymnaea stagnalis, and Helisoma trivolvis. Eight of the mesocosms received 45 Bellamya, eight received a pair of crayfish (4), eight received both Bellamya and crayfish, and eight were held as controls.
The most striking result of these experiments was that the populations of Physa and Lymnaea grew in the absence of Bellamya, but declined in the presence. The control Physa roughly doubled in population size over the 8 weeks, while the control Lymnaea roughly doubled in wet mass, although apparently did not reproduce (5). But all pulmonate populations declined by all measures in the Bellamya treatments. Clearly the invasion of Bellamya through the lakes of Northern Wisconsin should have negative effects on the native freshwater gastropods, right?Well, of course it’s more complicated than that. The Helisoma populations declined in both biomass and abundance (5) in all mesocosms, including the controls. Whether the Helisoma were outcompeted by the Physa and Lymnaea, or whether these mesocosms were simply unsuitable habitat for Helisoma, we'll never know. My intuition suggests to me that if the researchers had floated some macrophytic vegetation on the surface of their mesocosms, their Helisoma populations would have been fine (6). The problem is that the experiments of Johnson and colleagues show Helisoma going to extinction in the absence of Bellamya just as clearly as they show Physa and Lymnaea going to extinction in the presence (7). As much as I love experiments, a 5’ x 5’ structural foam plastic tank and a lake are two entirely different things.
Who could disagree? Certainly not our colleagues in Wisconsin. Thus in a companion study, Chris Solomon and a gang of four (including yours truly) surveyed 42 lakes in northern Wisconsin, roughly half of which had been invaded by Bellamya and half not, collecting a great variety of environmental measures in the process (8).
We found evidence of positive correlations between the presence of Bellamya and several general measures of lake productivity, such as conductivity and Secchi turbidity, but very little evidence of an effect of Bellamya on the native freshwater gastropod fauna. At neither the site level nor at the lake level could we find any evidence that Bellamya presence or abundance had any effect on the native freshwater gastropod community at all.
The absence of any significant results whatsoever neatly confirms my hypothesis, of course. Casting an eye back through the FWGNA archives, it looks as though I’ve authored fully 13 previous posts on invasive species, primarily Pomacea, Potamopyrgus, and Bellamya, with other viviparids and Bithynia making cameo appearances. And it may be recalled that I am an advocate of the “empty niche” hypothesis, which dates me back to the 1970s with most embarrassing accuracy.
Essentially, I think that the two best predictors for the success of a potential invasion are that the invading population must be weedy and different (9). Invaders must be adapted to the new environment they are invading, of course, while ecologically different from the native inhabitants – the more different, the better.
The gigantic, filter-feeding, ovoviviparous Bellamya is indeed strikingly different from all the other elements of the freshwater gastropod community native to northern Wisconsin. Hence I would not expect to uncover any competitive effects, and feel quite validated that we did not find any.
But I hasten to add two asterisks. First, populations of four different species of viviparids inhabit the lakes of northern Wisconsin, three of which are gigantic, filter-feeding, ovoviviparous invaders: Bellamya chinensis, B. japonica, and Viviparus georgianus. That our distributional data returned no evidence of a negative relationship within this subset, over 197 sites in 42 lakes, does indeed seem to suggest that our methods may have been too weak to detect even bona fide competition, where it occurs.
Second, a special challenge attends the selection of any invasive species for a study of population biology. What would make the researcher think that any such population has reached the carrying capacity of its environment? Competition cannot begin until some resource becomes limiting. If both Bellamya and Viviparus populations are still growing and spreading through northern Wisconsin in 2009, there is no reason to imagine that either will have any effect on the other.
And in fact, our analysis did return evidence of correlations between Bellamya presence and such measures of disturbance such as boat landings and shoreline housing. This implies to us that Bellamya populations may indeed be actively growing and spreading even in northern Wisconsin, where the invasion may have been ongoing for as much as 50 years.
I conclude with a call for additional research (10). This is a button you have rarely seen me push in this series of essays, both because it is trivially obvious to me that we need additional research on all aspects of freshwater gastropod biology, and because I try not to preach to the choir.
But where other people see "invasive species," I see "model organism." We have been greatly enriched, for example, by a huge volume of first-rate research on Pomacea in recent years (11), and that literature is huge (12). Meanwhile I haven't seen more than a couple papers published on even the most basic aspects of the life history of Bellamya, in total, over the 100-year history of the North American invasion (13). And three paragraphs ago I found myself offering an hypothesis regarding carrying capacities of invasive viviparids without a single estimate of population size, even static, ever taken anytime, anywhere. As easy as invasive viviparids are to sample, sitting on clean sand at the bottom of bathtub reservoirs? Shame on us all!
Keep in touch, everybody!
Rob
Notes
(1) I follow Smith (2000) in preferring the (internationally-recognized) generic nomen "Bellamya" over the (provincial) "Cipangopaludina." See the FWGNA page on B. japonica for details and references.
(2) Wisconsin populations are almost entirely Bellamya chinensis. I did identify B. japonica in five (of 42) lakes surveyed, which were lumped with B. chinensis for analysis. That's B. chinensis on the left, and B. japonica on the right.

(3) Johnson, P. J., J. D. Olden, C. T. Solomon, and M. J. Vander Zanden (2009) Interactions among invaders: community and ecosystem effects of multiple invasive species in an experimental aquatic system. Oecologia 159: 161-170.
(4) The crayfish were the invasive Orconectes rusticus. Those results were interesting as well, but don’t bear directly on the subject of this essay.
(5) Pulmonate juveniles are born at approximately 1 mm shell length. The sampling problems of finding such “speck babies” in a 1,000 liter mesocosm with sand and cinder blocks should be obvious to everybody. Thus I strongly suspect the “pulmonate abundance” figures reported by Johnson and colleagues are systematic underestimates.
(6) Helisoma is certainly quite commonly observed on floating vegetation. Any sort of additional habitat along these lines would certainly have benefited all three pulmonate populations.
(7) And if there’s a crayfish in the lake, all pulmonate populations are immediately doomed.
(8) Solomon, C. T, J. D. Olden, P. T. J. Johnson, R. T. Dillon and M J. Vander Zanden (2010) Distribution and community-level effects of the Chinese mystery snail (Bellamya chinensis) in northern Wisconsin lakes. Biological Invasions 12: 1591 - 1605. [PDF] 476kb.
(9) "Invaders Great and Small" Post of September '08.
(10) This should not be construed as a call to sequence two genes from single individuals sampled from 40 imaginary viviparid species in a vain attempt to reconstruct evolutionary events of 100 million years ago, while remaining arrogantly clueless regarding the basic population genetics of even one crappy pond full of crappy snails today.
(11) "Ampullariids star at Asilomar" Post of August '05.
(12) "Review: Global Advances in Golden Apple Snails" Post of May '07.
(13) Stanczykowska, A, E. Magnin and A. Dumouchel (1971) Etude de trois populations de Viviparus malleatus (Reeve) de la region de Montreal. I. Croissance, fecondite, biomasse et production annuelle. Can. J. Zool. 49: 1431-1441. Jokinen, E.H. (1982) Cipangopaludina chinensis (Gastropoda: Viviparidae) in North America, review and update. Nautilus 96: 89-95. Therriault, T. W. and E. Kott (2002-3) Cipangopaludina chinensis malleata (Gastropoda: Viviparidae) in southern Ontario: An update of the distribution and some aspects of life history. Malac. Rev. 35-36: 111-121.
Comments
(1) From John Havel JohnHavel@MissouriState.edu
Hi Rob,
Since I’m a newcomer to snail research (although an old hand at invasions), I was hesitant to reply to all. So, below are my comments on your most-interesting essay. I plan to work again next spring and summer at Trout Lake. Doing some quantitative sampling for densities and fecundity patterns could be interesting. Care to join me for part of the venture?
Nice review of the papers by Johnson et al. (2009) and Solomon et al (in press). By the way, in last summer’s experiments with a former student, I also found very little effect of Bellamya on Physa growth rates in the laboratory (Clark and Havel, unpublished data).
See also the paper of Pattinson et al. (2003) on invasive Daphnia. We discuss the same empty niche hypothesis as important for tropical D. lumholtzi invading reservoirs of the south-central US.
I don’t agree that Bellamya are that different in dominant feeding ecology from native gastropods. After all, Bellamya are well known to clean surfaces quite thoroughly, and are for sale by breeders on the internet for cleaning ornamental ponds. Perhaps more important is whether or not resources are indeed limiting. (Although I now see that you consider this later in your essay.)
Regarding your statement about Bellamya being a "model organism." I agree! Lots of ways to study general ecology principles. [e.g., see papers by Havel and Shurin (2004) and Havel et al. (2005)]
Your assertion that we are "without a single estimate of population size, even static, ever taken anytime, anywhere" is an exaggeration, of course. See Solomon et al. (2009): quantitative sampling (see methods par. 4) plus population density estimates (results par. 5). But, I agree that population and fecundity estimates would be useful for better understanding these study systems and the potential of exotic viviparid snails for population growth and expansion.
But regarding your suggestion that viviparids might be easy to sample, "sitting on clean sand at the bottom of bathtub reservoirs," lakes can be spatially very complex (macrophytes, rocks, sand, logs, leaf litter, flocculent organics). I found quantitative sampling to be a real challenge; but then I’m new to the benthos. (I could definitely use some help with these techniques.)
Additional references
Havel, J.E., and J.B. Shurin. 2004. Mechanisms, effects, and scales of dispersal in freshwater zooplankton: a synthesis. Limnology and Oceanography 49: 1229-1238. Havel, J.E., C. E. Lee, and J. Vander Zanden. 2005. Do reservoirs facilitate invasions into landscapes? BioScience 55: 518-525. Pattinson, K.R., J.E. Havel, and R.G. Rhodes. 2003. Invasibility of a reservoir to exotic Daphnia lumholtzi: Experimental assessment of diet selection and life history responses to Cyanobacteria. Freshwater Biology 48: 233-246.
(2) From David Campbell amblema@bama.ua.edu
Regarding Smith's (2000) preference for the generic nomen "Bellamya," unfortunately, he's led you astray. The type of Bellamya is from West Africa, and no East Asian species belongs in it (even if you put them in the same genus, there are older names both from Africa and from Asia). See... Mita E. Sengupta, Thomas K. Kristensen, Henry Madsen, Aslak Jørgensen. 2009. Molecular phylogenetic investigations of the Viviparidae (Gastropoda: Caenogastropoda) in the lakes of the Rift Valley area of Africa. Molecular Phylogenetics and Evolution 52:797–805 ...for the molecular data, though they don't deal with the nomenclatural implications, except for incorrectly suggesting that Neothauma should be included in Bellamya- Neothauma is the older name. It's still possible that some other name applies besides Cipangpaludina, but to determine that requires actually checking the type species of the nominal Asian genera.
(3) From Jeremy Tiemann jtiemann@inhs.uiuc.edu
Have you seen this paper yet? Bury, J.A. B.E. Sietman, and B.N. Karns. 2007. Distribution of the non-native viviparid snails, Bellamya chinensis and Viviparus georgianus, in Minnesota and the first record of Bellamya japonica from Wisconsin. Journal of Freshwater Ecology 22(4):697-703.
(4) From Bob Prezant prezantr@mail.montclair.edu
Rob, I agree...a "model organism" is just what we have in Bellamya. Can't remember if I sent this article to you. Hope you're warmer than we are here in NJ. Prezant, R. S., E. J. Chapman and A. McDougall (2006) In utero predator-induced responses in the viviparid snail Bellamya chinensis. Canadian Journal of Zoology 84: 600-608.
Friday, November 13, 2009
Mobile Basin IV: Goniobasis WTFs
Editor's Note. This essay was subsequently published as: Dillon, R.T., Jr. (2019c) Mobile Basin III: Goniobasis clues. Pp. 35-39 in The Freshwater Gastropods of North America Volume 3, Essays on the Prosobranchs. FWGNA Press, Charleston.
Georgia is just Florida, one state north and 65 million years back. During the Cretaceous Period, most of the land mass that constitutes the present Mobile Basin of Alabama would have been obscured by a shallow sea and the occasional shadow of a passing mosasaur. The exception would have been a peninsula in what is today North Georgia (1).
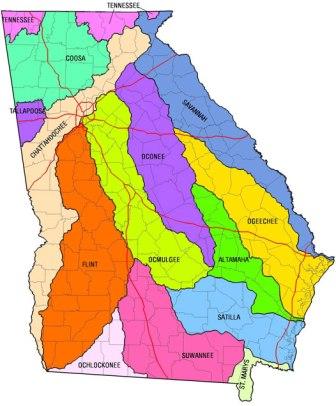
Atlanta sits in the middle of that Cretaceous peninsula, General Sherman notwithstanding (2). The low hills to its east drain through the Ocmulgee/Oconee/Altamaha to the Atlantic. The Chattahoochee River runs through the middle of the city, under the I-285 beltway and southwest to the Gulf. To its immediate north lie tributaries of the Alabama/Coosa, draining into the Mobile Basin. And just a bit further north, the waters of the Tennessee River flow into the Mississippi (below, 3).
 An appreciation of the complex drainage patterns of north Georgia will be important for an understanding of the hypotheses I am preparing to advance in this essay. If you are planning an earnest attempt to follow me, you will need to hold in your mind that the Coosa River is formed at the Georgia/Alabama line by the junction of two major tributaries, the (more northern) Conasauga/ Coosawattee/ Oostanaula and the (more southern) Etowah (4). And I do apologize for the drainage taxonomy. Subsequent to the departure of the mosasaurs, but prior to the arrival of Sherman, this part of the world was infested with dozens of Indian tribes, vying among one another in the unpronouncability of their names. After extinguishing each tribe, the European settlers named a local river in its honor, apparently careless of how, or even whether, any of these rivers might ultimately interconnect.
An appreciation of the complex drainage patterns of north Georgia will be important for an understanding of the hypotheses I am preparing to advance in this essay. If you are planning an earnest attempt to follow me, you will need to hold in your mind that the Coosa River is formed at the Georgia/Alabama line by the junction of two major tributaries, the (more northern) Conasauga/ Coosawattee/ Oostanaula and the (more southern) Etowah (4). And I do apologize for the drainage taxonomy. Subsequent to the departure of the mosasaurs, but prior to the arrival of Sherman, this part of the world was infested with dozens of Indian tribes, vying among one another in the unpronouncability of their names. After extinguishing each tribe, the European settlers named a local river in its honor, apparently careless of how, or even whether, any of these rivers might ultimately interconnect.
In any case, the diversity of the Goniobasis fauna of the Alabama/Coosa river system might aptly be described as "legendary." Goodrich (1936) recognized 24 species in the main river itself, and subsequently (1941) added about 11 species from "small streams." His final (1944) tally for the entire fauna of the Coosa River system listed, from rivers of all size, 29 species of Goniobasis, many with subspecies (5).
As patient and perceptive readers of this series of essays may have by now gathered, I myself have relatively little field experience anywhere in the Mobile Basin. What insight I have gathered regarding the pleurocerid fauna of the Oostanaula side of the Coosa has developed as a consequence of my work in east Tennessee on Goniobasis clavaeformis / Pleurocera unciale, as featured in last month’s (6) “Mobile Basin III.” When first I crossed the low hills that separate the Hiwassee system of the Tennessee River from the Mobile Basin in North Georgia, and peeked tentatively into the famously diverse waters of the Oostanaula subdrainage, I was shocked by the sight that met my eyes. WTF?!
This was the same pleurocerid fauna I had just spent the previous week sampling in Tennessee. In Cohulla Creek, one of the main tributaries of the Conasauga/Oostanaula east of Dalton, I recognized three old friends: Goniobasis simplex, G. troostiana (or "arachnoidea"), and G. clavaeformis – the same species widespread from southwest Virginia down the width of east Tennessee. The only difference was that clavaeformis was now called vestita, arachnoidea was now called striatula, and simplex was now called, well, I don't know (7). And the allozyme data I have subsequently collected seem to show that I was correct.
But it had happened before. My first “WTF moment” in the upper Mobile Basin actually occurred in the mid-1990s, kneeling beside the Etowah River. On that occasion I had been sampling west from the Atlantic drainages of central Georgia, crossed the broken Piedmont north of Atlanta, and found essentially the same G. catenaria I’d I had spent most of my professional career chasing through southern Virginia and the Carolinas, now re-named “Goniobasis caelatura” in the Etowah. And the DNA sequence data I have subsequently gathered with the help of my colleague John Robinson again seem to show that this impression was correct (8).
So we’ve now reached the point of a hypothesis – not just for this essay, but for all IV essays in this series. The endemicity of the Mobile Basin pleurocerid fauna has historically been much overstated. As the Cretaceous seas receded, the pleurocerid faunas that colonized the Mobile Basin were a mixture of the Atlantic (older) elements and the Tennessee (younger) elements, both previously inhabiting a peninsula in north Georgia. These faunas can still be seen today in the two major tributaries of the Coosa – the southern Etowah and the northern Oostanaula.
But don’t misunderstand me. There are bona fide endemic species of pleurocerid gastropods in the Mobile Basin. Dykes Creek is a lovely little stream running directly into the main Etowah River very near its junction with the Oostanaula. It is inhabited by (at least) three species: a G. clavaeformis-type from the Tennessee side (A), a chunky G. catenaria from the Atlantic side (B), and a species I'm going to call "WTF3" (C). Allozyme data show WTF3 to be genetically similar to G. catenaria, but reproductively isolated from it. My best guess is that Goodrich would have identified WTF3 as the subspecies Goniobasis caelatura sternsiana (Call 1886). But if so, that's the second of his caelatura "subspecies" that needs to be raised back up to the full species level (9). I am quite sure, in any case, that WTF3 is endemic to the Mobile Basin. And perhaps very rare.
 So to summarize. In late June the US Fish and Wildlife service proposed endangered status for two Mobile Basin pleurocerids species, "Leptoxis foremani" and "Pleurocera foremani" (Essay I, 24Aug 09 - see note 10). The former nominal species is likely a local population of L. picta, which is not protected in any way (Essay II, 15Sept - see note 11). The latter nominal species is likely a local ecophenotypic variant of the most widespread pleurocerid in the American southeast (Essay III, 12Oct - see note 6). Meanwhile genuinely endemic and probably rare species go entirely unrecognized by anybody.
So to summarize. In late June the US Fish and Wildlife service proposed endangered status for two Mobile Basin pleurocerids species, "Leptoxis foremani" and "Pleurocera foremani" (Essay I, 24Aug 09 - see note 10). The former nominal species is likely a local population of L. picta, which is not protected in any way (Essay II, 15Sept - see note 11). The latter nominal species is likely a local ecophenotypic variant of the most widespread pleurocerid in the American southeast (Essay III, 12Oct - see note 6). Meanwhile genuinely endemic and probably rare species go entirely unrecognized by anybody.
Science and public policy are two entirely different things. What I have been demonstrating over this four part series is science - the construction of testable hypotheses about the natural world. This process, this vocabulary, this way of looking at the world, is entirely different from the process that brought the US Fish & Wildlife Service to propose its endangered species regulations in the 29Jun09 Federal Register. These two ways of looking at the world are not compatible, nor are they incompatible. FWS biologists play baseball, I play the banjo. Neither is better, neither is worse.
I am not suggesting that the FWS drop "Leptoxis foremani" and "Pleurocera foremani" like a pair of old girlfriends and drive Dad's Chrysler up to Dykes Creek with flowers and bonbons to court G. stearnsiana, or whatever TF it is. Everything I've offered in this essay is a hypothesis (12), and may change tomorrow. What I am suggesting is that if it is the genuine Will of the People that public resources be expended on science (13), all government agencies involved (regardless of mission) must support what has heretofore been marginalized as "basic research," such as that which produced the hypotheses I have advanced above, which is, after all, the only real Science there is. In other words - send me money, and leave me TF alone.
I'm waiting,
Rob
Notes
(1) The best treatment of this subject I've been able to find on the web is at NOAA, interestingly: Beach Nourishment, A Guide for Local Government Officials
(2) Literally.
(3) I've stolen this map from the "Southeast Watershed Assistance Network."
(4) And here's the map of the Mobile Basin drainage system that I stole for Essay I in this series. You might want to open it in a new window.
(5) Goodrich, C. (1936) Goniobasis of the Coosa River, Alabama. Misc. Publ. Mus. Zool. Univ. Mich., 31, 1-60. Goodrich, C. (1941) Pleuroceridae of the small streams of the Alabama River system. Occas. Pprs. Mus. Zool. Univ. Mich., 427, 1-10. Goodrich, C. (1944) Pleuroceridae of the Coosa River basin. Nautilus, 58, 40-48.
(6) "Mobile Basin III: Pleurocera Puzzles." Post of October '09.
(7) This is actually one of the biggest mysteries in my mind regarding the Goniobasis of the Alabama/Coosa. What did Goodrich (and everybody else who has ever sloshed through these rivers) call plain old, ordinary, vanilla, dirt-common Goniobasis simplex? I can't match it to a synonym on anybody's list, over 150 years of surveys. Has it been missed?
(8) "The Snails the Dinosaurs Saw." Post of March '09.
(9) In the mid-1990s I also recognized Goniobasis georgiana in the Etowah, another older-Appalachian species ranging through the upper tributaries of the Hiwassee in the North Carolina mountains. Goodrich lowered this nomen to subspecific rank as "G. caelatura georgiana," but its co-occurrence in the Etowah with what (I suppose) he would have identified as G. caelatura caelatura, absent any evidence of hybridization, suggests strongly that G. georgiana is a distinct species.
(10) "Mobile Basin I: Two pleurocerids proposed for listing." Post of August '09.
(11) "Mobile Basin II: Leptoxis lessons." Post of September '09.
(12) Thompson (2000, Walkerana 11:1-54) has a different hypothesis. In the words of my favorite Alabamian, "That's all I've got to say about that."
(13) And I'm not sure it is!
Georgia is just Florida, one state north and 65 million years back. During the Cretaceous Period, most of the land mass that constitutes the present Mobile Basin of Alabama would have been obscured by a shallow sea and the occasional shadow of a passing mosasaur. The exception would have been a peninsula in what is today North Georgia (1).
Atlanta sits in the middle of that Cretaceous peninsula, General Sherman notwithstanding (2). The low hills to its east drain through the Ocmulgee/Oconee/Altamaha to the Atlantic. The Chattahoochee River runs through the middle of the city, under the I-285 beltway and southwest to the Gulf. To its immediate north lie tributaries of the Alabama/Coosa, draining into the Mobile Basin. And just a bit further north, the waters of the Tennessee River flow into the Mississippi (below, 3).
 An appreciation of the complex drainage patterns of north Georgia will be important for an understanding of the hypotheses I am preparing to advance in this essay. If you are planning an earnest attempt to follow me, you will need to hold in your mind that the Coosa River is formed at the Georgia/Alabama line by the junction of two major tributaries, the (more northern) Conasauga/ Coosawattee/ Oostanaula and the (more southern) Etowah (4). And I do apologize for the drainage taxonomy. Subsequent to the departure of the mosasaurs, but prior to the arrival of Sherman, this part of the world was infested with dozens of Indian tribes, vying among one another in the unpronouncability of their names. After extinguishing each tribe, the European settlers named a local river in its honor, apparently careless of how, or even whether, any of these rivers might ultimately interconnect.
An appreciation of the complex drainage patterns of north Georgia will be important for an understanding of the hypotheses I am preparing to advance in this essay. If you are planning an earnest attempt to follow me, you will need to hold in your mind that the Coosa River is formed at the Georgia/Alabama line by the junction of two major tributaries, the (more northern) Conasauga/ Coosawattee/ Oostanaula and the (more southern) Etowah (4). And I do apologize for the drainage taxonomy. Subsequent to the departure of the mosasaurs, but prior to the arrival of Sherman, this part of the world was infested with dozens of Indian tribes, vying among one another in the unpronouncability of their names. After extinguishing each tribe, the European settlers named a local river in its honor, apparently careless of how, or even whether, any of these rivers might ultimately interconnect.In any case, the diversity of the Goniobasis fauna of the Alabama/Coosa river system might aptly be described as "legendary." Goodrich (1936) recognized 24 species in the main river itself, and subsequently (1941) added about 11 species from "small streams." His final (1944) tally for the entire fauna of the Coosa River system listed, from rivers of all size, 29 species of Goniobasis, many with subspecies (5).
As patient and perceptive readers of this series of essays may have by now gathered, I myself have relatively little field experience anywhere in the Mobile Basin. What insight I have gathered regarding the pleurocerid fauna of the Oostanaula side of the Coosa has developed as a consequence of my work in east Tennessee on Goniobasis clavaeformis / Pleurocera unciale, as featured in last month’s (6) “Mobile Basin III.” When first I crossed the low hills that separate the Hiwassee system of the Tennessee River from the Mobile Basin in North Georgia, and peeked tentatively into the famously diverse waters of the Oostanaula subdrainage, I was shocked by the sight that met my eyes. WTF?!
This was the same pleurocerid fauna I had just spent the previous week sampling in Tennessee. In Cohulla Creek, one of the main tributaries of the Conasauga/Oostanaula east of Dalton, I recognized three old friends: Goniobasis simplex, G. troostiana (or "arachnoidea"), and G. clavaeformis – the same species widespread from southwest Virginia down the width of east Tennessee. The only difference was that clavaeformis was now called vestita, arachnoidea was now called striatula, and simplex was now called, well, I don't know (7). And the allozyme data I have subsequently collected seem to show that I was correct.
But it had happened before. My first “WTF moment” in the upper Mobile Basin actually occurred in the mid-1990s, kneeling beside the Etowah River. On that occasion I had been sampling west from the Atlantic drainages of central Georgia, crossed the broken Piedmont north of Atlanta, and found essentially the same G. catenaria I’d I had spent most of my professional career chasing through southern Virginia and the Carolinas, now re-named “Goniobasis caelatura” in the Etowah. And the DNA sequence data I have subsequently gathered with the help of my colleague John Robinson again seem to show that this impression was correct (8).
So we’ve now reached the point of a hypothesis – not just for this essay, but for all IV essays in this series. The endemicity of the Mobile Basin pleurocerid fauna has historically been much overstated. As the Cretaceous seas receded, the pleurocerid faunas that colonized the Mobile Basin were a mixture of the Atlantic (older) elements and the Tennessee (younger) elements, both previously inhabiting a peninsula in north Georgia. These faunas can still be seen today in the two major tributaries of the Coosa – the southern Etowah and the northern Oostanaula.
But don’t misunderstand me. There are bona fide endemic species of pleurocerid gastropods in the Mobile Basin. Dykes Creek is a lovely little stream running directly into the main Etowah River very near its junction with the Oostanaula. It is inhabited by (at least) three species: a G. clavaeformis-type from the Tennessee side (A), a chunky G. catenaria from the Atlantic side (B), and a species I'm going to call "WTF3" (C). Allozyme data show WTF3 to be genetically similar to G. catenaria, but reproductively isolated from it. My best guess is that Goodrich would have identified WTF3 as the subspecies Goniobasis caelatura sternsiana (Call 1886). But if so, that's the second of his caelatura "subspecies" that needs to be raised back up to the full species level (9). I am quite sure, in any case, that WTF3 is endemic to the Mobile Basin. And perhaps very rare.
So to summarize. In late June the US Fish and Wildlife service proposed endangered status for two Mobile Basin pleurocerids species, "Leptoxis foremani" and "Pleurocera foremani" (Essay I, 24Aug 09 - see note 10). The former nominal species is likely a local population of L. picta, which is not protected in any way (Essay II, 15Sept - see note 11). The latter nominal species is likely a local ecophenotypic variant of the most widespread pleurocerid in the American southeast (Essay III, 12Oct - see note 6). Meanwhile genuinely endemic and probably rare species go entirely unrecognized by anybody.Science and public policy are two entirely different things. What I have been demonstrating over this four part series is science - the construction of testable hypotheses about the natural world. This process, this vocabulary, this way of looking at the world, is entirely different from the process that brought the US Fish & Wildlife Service to propose its endangered species regulations in the 29Jun09 Federal Register. These two ways of looking at the world are not compatible, nor are they incompatible. FWS biologists play baseball, I play the banjo. Neither is better, neither is worse.
I am not suggesting that the FWS drop "Leptoxis foremani" and "Pleurocera foremani" like a pair of old girlfriends and drive Dad's Chrysler up to Dykes Creek with flowers and bonbons to court G. stearnsiana, or whatever TF it is. Everything I've offered in this essay is a hypothesis (12), and may change tomorrow. What I am suggesting is that if it is the genuine Will of the People that public resources be expended on science (13), all government agencies involved (regardless of mission) must support what has heretofore been marginalized as "basic research," such as that which produced the hypotheses I have advanced above, which is, after all, the only real Science there is. In other words - send me money, and leave me TF alone.
I'm waiting,
Rob
Notes
(1) The best treatment of this subject I've been able to find on the web is at NOAA, interestingly: Beach Nourishment, A Guide for Local Government Officials
(2) Literally.
(3) I've stolen this map from the "Southeast Watershed Assistance Network."
(4) And here's the map of the Mobile Basin drainage system that I stole for Essay I in this series. You might want to open it in a new window.
(5) Goodrich, C. (1936) Goniobasis of the Coosa River, Alabama. Misc. Publ. Mus. Zool. Univ. Mich., 31, 1-60. Goodrich, C. (1941) Pleuroceridae of the small streams of the Alabama River system. Occas. Pprs. Mus. Zool. Univ. Mich., 427, 1-10. Goodrich, C. (1944) Pleuroceridae of the Coosa River basin. Nautilus, 58, 40-48.
(6) "Mobile Basin III: Pleurocera Puzzles." Post of October '09.
(7) This is actually one of the biggest mysteries in my mind regarding the Goniobasis of the Alabama/Coosa. What did Goodrich (and everybody else who has ever sloshed through these rivers) call plain old, ordinary, vanilla, dirt-common Goniobasis simplex? I can't match it to a synonym on anybody's list, over 150 years of surveys. Has it been missed?
(8) "The Snails the Dinosaurs Saw." Post of March '09.
(9) In the mid-1990s I also recognized Goniobasis georgiana in the Etowah, another older-Appalachian species ranging through the upper tributaries of the Hiwassee in the North Carolina mountains. Goodrich lowered this nomen to subspecific rank as "G. caelatura georgiana," but its co-occurrence in the Etowah with what (I suppose) he would have identified as G. caelatura caelatura, absent any evidence of hybridization, suggests strongly that G. georgiana is a distinct species.
(10) "Mobile Basin I: Two pleurocerids proposed for listing." Post of August '09.
(11) "Mobile Basin II: Leptoxis lessons." Post of September '09.
(12) Thompson (2000, Walkerana 11:1-54) has a different hypothesis. In the words of my favorite Alabamian, "That's all I've got to say about that."
(13) And I'm not sure it is!
Monday, October 12, 2009
Mobile Basin III: Pleurocera Puzzles
Editor's Note - The research results telegraphed below were ultimately published by Dillon (2011) in Malacologia 53: 265 - 277.
Last month (1) we made passing reference to the Awakening of Calvin Goodrich, that 1934-41 period during which he published “Studies of the Gastropod Family Pleuroceridae” I – VIII (2). During this stage of his career, our hero developed his thesis that the characters upon which most pleurocerid taxonomy had been based - shell dimensions, coloration, ornamentation and so forth - are highly plastic and subject to environmental influence (3). In 2007 I coined the term “Goodrichian taxon shift” to describe situations where a single population of pleurocerids might vary down an environmental gradient to such an extent that taxonomists have recognized two different species (4). Or perhaps even two different genera?
I realize that the heading at the top of this essay is “Mobile Basin III.” But I beg your indulgence for the next three paragraphs to stray one drainage north, to the headwaters of the Tennessee River.
In my "Goodrichian" essay of 20Feb07 I reported unpublished observations on variation at two allozyme loci in a population of Goniobasis from Indian Creek, a tributary of the Clinch/Powell in southwest Virginia (5). In the headwaters, this single population of pleurocerids bore shells that were strongly carinate (historically referred to "G. acutocarinata" CA1), in middle reaches their shell morphology was smooth and typical of G. clavaeformis (C1), and upon joining the main Powell River, their shells became chunky and angulate, showing the morphology generally assigned the nomen “Pleurocera unciale” (P1).
In the last couple years I have extended this research program down the width of East Tennessee, to include populations from the Little River drainage near Maryville (figs CA2, C2, P2 at left), the Conasauga/Hiwassee near Etowah (figs CA3, C6, P3), and the Coahulla/Oostanaula/Coosa in North Georgia (figs CA4, P4). I have also sampled populations of G. simplex from each of these four regions to calibrate expected levels of interpopulation divergence at the 10 allozyme loci examined (fig S7).
The results of my 2007 study in Indian Creek are confirmed (6). Each of the four samples of “Pleurocera” was more genetically similar to the Goniobasis population just upstream than to any other sample of Pleurocera. Apparently taxonomists have been misled by ecophenotypic variation to identify populations of a single, widespread species under two different genera.
This result extends from the rivers of Tennessee into the Coosa drainage of North Georgia. Tradition has always held that elements of the Mobile Basin fauna are treated as endemic, even when indistinguishable from the fauna of neighboring drainages. Thus Goniobasis populations from small creeks in the Alabama/Coosa drainage showing the strongly carinate shell morphology are typically identified as “Goniobasis carinifera,” and those with heavier, smoother shells downstream as “Pleurocera vestitum.” Interestingly, if carinifera/vestitum is indeed conspecific with clavaeformis/unciale, the oldest name for the entire, sprawling species, from Virginia to Alabama, would be the Coosa name Goniobasis carinifera (Lamarck 1822).
In August we kicked this series off with news that the US Fish & Wildlife Service has proposed Alabama populations of pleurocerids identified as “Pleurocera foremani” for listing under the Endangered Species Act (7). According to the notice posted in the Federal Register, P. foremani is found today at only two locations, the lower Coosa River below Wetumpka Shoals and lower Yellowleaf Creek, a tributary of the Coosa (8). I understand from colleagues with field experience in this region that, sampling upstream, P. foremani is replaced by Pleurocera prasinatum in the smaller rivers, and that P. prasinatum is replaced by Pleurocera vestitum, and that vestitum is replaced by Goniobasis carinifera in the creeks.
In 1944, the mature Calvin Goodrich wrote regarding the Coosa species of Pleurocera, "On close study and comparison, they resolve themselves into four forms, and even these are not very distinctive. The specific names, in short, are to be considered conveniences in sorting rather than clean-cut differentiations" (9). He then went on to list P. vestitum "especially common in head streams," P. prasinatum "in the middle and lower Coosa," and P. foremani, which he distinguished by shell sculpture that "in one locality of the Cahaba is plainly a reversion, the same thing may be true of the shells of the Coosa.
"I have not seen the dissertation of Jeffrey Sides. But again referring to the 29Jun09 Federal Register (8), his sequence data suggested that P. foremani "was genetically more closely allied to a co-occurring species in the genus Elimia (Goniobasis)" than to any other species in the genus Pleurocera. Really? Even though double-digit sequence divergence is not uncommon within even conspecific populations of pleurocerids (10), the divergence between P. foremani and the local upstream Goniobasis appears to be of minor consequence?
Is the foremani/prasinatum/vestitum/carinifera taxon continuum in Alabama nothing but a unciale/clavaeformis/acutocarinata Goodrichian taxon shift, one state south? Has the US Fish and Wildlife service proposed federal protection for a local ecophenotypic variant of the most widespread pleurocerid in the American southeast? Pleurocera puzzles, indeed!
Stay tuned for more ...
Rob
Notes
(1) "Mobile Basin II: Leptoxis Lessons." Post of September '09 .
(2) A nice Goodrich bibliography is available from Kevin Cummings' website at the INHS
(3) "The Legacy of Calvin Goodrich." Post of January '07.
(4) "Goodrichian Taxon Shift." Post of February '07.
(5) Dillon, R.T. & J. D. Robinson (2007) The Goniobasis ("Elimia") of southwest Virginia, II. Shell morphological variation in Goniobasis clavaeformis. Report to the Virginia Division of Game and Inland Fisheries, contract 2006-9308. 12 pp. [pdf]
(6) I presented this research at a NABS symposium this May in Grand Rapids. A manuscript is currently in preparation. Dillon, R. T. (in prep) Genetic and morphological divergence among populations of pleurocerid snails inhabiting rivers of the Southern Appalachians: Evidence of a two-stage process.
(7) "Mobile Basin I: Two Pleurocerids Proposed for Listing." Post of August '09.
(8) Follow the link from the FWS Press Release: Service Proposes Endangered Species Status and Critical Habitat Designations for the Georgia Pigtoe Mussel, Interrupted Rocksnail, and Rough Hornsnail.
(9) Goodrich, C. 1944. Pleuroceridae of the Coosa River basin. Nautilus 58(2):40-48.
(10) "The Snails the Dinosaurs Saw." Post of March '09.
Last month (1) we made passing reference to the Awakening of Calvin Goodrich, that 1934-41 period during which he published “Studies of the Gastropod Family Pleuroceridae” I – VIII (2). During this stage of his career, our hero developed his thesis that the characters upon which most pleurocerid taxonomy had been based - shell dimensions, coloration, ornamentation and so forth - are highly plastic and subject to environmental influence (3). In 2007 I coined the term “Goodrichian taxon shift” to describe situations where a single population of pleurocerids might vary down an environmental gradient to such an extent that taxonomists have recognized two different species (4). Or perhaps even two different genera?
I realize that the heading at the top of this essay is “Mobile Basin III.” But I beg your indulgence for the next three paragraphs to stray one drainage north, to the headwaters of the Tennessee River.

In my "Goodrichian" essay of 20Feb07 I reported unpublished observations on variation at two allozyme loci in a population of Goniobasis from Indian Creek, a tributary of the Clinch/Powell in southwest Virginia (5). In the headwaters, this single population of pleurocerids bore shells that were strongly carinate (historically referred to "G. acutocarinata" CA1), in middle reaches their shell morphology was smooth and typical of G. clavaeformis (C1), and upon joining the main Powell River, their shells became chunky and angulate, showing the morphology generally assigned the nomen “Pleurocera unciale” (P1).
In the last couple years I have extended this research program down the width of East Tennessee, to include populations from the Little River drainage near Maryville (figs CA2, C2, P2 at left), the Conasauga/Hiwassee near Etowah (figs CA3, C6, P3), and the Coahulla/Oostanaula/Coosa in North Georgia (figs CA4, P4). I have also sampled populations of G. simplex from each of these four regions to calibrate expected levels of interpopulation divergence at the 10 allozyme loci examined (fig S7).
The results of my 2007 study in Indian Creek are confirmed (6). Each of the four samples of “Pleurocera” was more genetically similar to the Goniobasis population just upstream than to any other sample of Pleurocera. Apparently taxonomists have been misled by ecophenotypic variation to identify populations of a single, widespread species under two different genera.
This result extends from the rivers of Tennessee into the Coosa drainage of North Georgia. Tradition has always held that elements of the Mobile Basin fauna are treated as endemic, even when indistinguishable from the fauna of neighboring drainages. Thus Goniobasis populations from small creeks in the Alabama/Coosa drainage showing the strongly carinate shell morphology are typically identified as “Goniobasis carinifera,” and those with heavier, smoother shells downstream as “Pleurocera vestitum.” Interestingly, if carinifera/vestitum is indeed conspecific with clavaeformis/unciale, the oldest name for the entire, sprawling species, from Virginia to Alabama, would be the Coosa name Goniobasis carinifera (Lamarck 1822).
In August we kicked this series off with news that the US Fish & Wildlife Service has proposed Alabama populations of pleurocerids identified as “Pleurocera foremani” for listing under the Endangered Species Act (7). According to the notice posted in the Federal Register, P. foremani is found today at only two locations, the lower Coosa River below Wetumpka Shoals and lower Yellowleaf Creek, a tributary of the Coosa (8). I understand from colleagues with field experience in this region that, sampling upstream, P. foremani is replaced by Pleurocera prasinatum in the smaller rivers, and that P. prasinatum is replaced by Pleurocera vestitum, and that vestitum is replaced by Goniobasis carinifera in the creeks.
In 1944, the mature Calvin Goodrich wrote regarding the Coosa species of Pleurocera, "On close study and comparison, they resolve themselves into four forms, and even these are not very distinctive. The specific names, in short, are to be considered conveniences in sorting rather than clean-cut differentiations" (9). He then went on to list P. vestitum "especially common in head streams," P. prasinatum "in the middle and lower Coosa," and P. foremani, which he distinguished by shell sculpture that "in one locality of the Cahaba is plainly a reversion, the same thing may be true of the shells of the Coosa.
"I have not seen the dissertation of Jeffrey Sides. But again referring to the 29Jun09 Federal Register (8), his sequence data suggested that P. foremani "was genetically more closely allied to a co-occurring species in the genus Elimia (Goniobasis)" than to any other species in the genus Pleurocera. Really? Even though double-digit sequence divergence is not uncommon within even conspecific populations of pleurocerids (10), the divergence between P. foremani and the local upstream Goniobasis appears to be of minor consequence?
Is the foremani/prasinatum/vestitum/carinifera taxon continuum in Alabama nothing but a unciale/clavaeformis/acutocarinata Goodrichian taxon shift, one state south? Has the US Fish and Wildlife service proposed federal protection for a local ecophenotypic variant of the most widespread pleurocerid in the American southeast? Pleurocera puzzles, indeed!
Stay tuned for more ...
Rob
Notes
(1) "Mobile Basin II: Leptoxis Lessons." Post of September '09 .
(2) A nice Goodrich bibliography is available from Kevin Cummings' website at the INHS
(3) "The Legacy of Calvin Goodrich." Post of January '07.
(4) "Goodrichian Taxon Shift." Post of February '07.
(5) Dillon, R.T. & J. D. Robinson (2007) The Goniobasis ("Elimia") of southwest Virginia, II. Shell morphological variation in Goniobasis clavaeformis. Report to the Virginia Division of Game and Inland Fisheries, contract 2006-9308. 12 pp. [pdf]
(6) I presented this research at a NABS symposium this May in Grand Rapids. A manuscript is currently in preparation. Dillon, R. T. (in prep) Genetic and morphological divergence among populations of pleurocerid snails inhabiting rivers of the Southern Appalachians: Evidence of a two-stage process.
(7) "Mobile Basin I: Two Pleurocerids Proposed for Listing." Post of August '09.
(8) Follow the link from the FWS Press Release: Service Proposes Endangered Species Status and Critical Habitat Designations for the Georgia Pigtoe Mussel, Interrupted Rocksnail, and Rough Hornsnail.
(9) Goodrich, C. 1944. Pleuroceridae of the Coosa River basin. Nautilus 58(2):40-48.
(10) "The Snails the Dinosaurs Saw." Post of March '09.
Tuesday, September 15, 2009
Mobile Basin II: Leptoxis Lessons
Editor's Note. This essay was subsequently published as: Dillon, R.T., Jr. (2019c) Mobile Basin II: Leptoxis legacy. Pp. 27-33 in The Freshwater Gastropods of North America Volume 3, Essays on the Prosobranchs. FWGNA Press, Charleston.
Through the early history of American malacology, the freshwater gastropod fauna of the Mobile Basin was not seen as exceptionally diverse. Isaac Lea, T. A. Conrad, and J. G. Anthony described about 20 species of Leptoxis (1) from the Mobile Basin from 1834 - 1860, for example, which Tryon (1873) boiled down to about 12 (2). This would not seem to be a significantly greater tally than the approximately 20 species of Leptoxis described from the Tennessee River system during that era.
 Some of my audience may recall that Calvin Goodrich began his career as a newspaperman, and that he was influenced by A. E. Ortmann to take up malacology around the years 1913 - 1917. One of Ortmann's closest professional colleagues was Herbert H. Smith, who retired in 1903 from the Carnegie Museum in Pittsburgh to become curator of the Alabama State Museum in Tuscaloosa. When H. H. Smith died in 1919, a review of Smith's collections of Alabama pleurocerids was passed by Ortmann to Goodrich.
Some of my audience may recall that Calvin Goodrich began his career as a newspaperman, and that he was influenced by A. E. Ortmann to take up malacology around the years 1913 - 1917. One of Ortmann's closest professional colleagues was Herbert H. Smith, who retired in 1903 from the Carnegie Museum in Pittsburgh to become curator of the Alabama State Museum in Tuscaloosa. When H. H. Smith died in 1919, a review of Smith's collections of Alabama pleurocerids was passed by Ortmann to Goodrich.
Through the early history of American malacology, the freshwater gastropod fauna of the Mobile Basin was not seen as exceptionally diverse. Isaac Lea, T. A. Conrad, and J. G. Anthony described about 20 species of Leptoxis (1) from the Mobile Basin from 1834 - 1860, for example, which Tryon (1873) boiled down to about 12 (2). This would not seem to be a significantly greater tally than the approximately 20 species of Leptoxis described from the Tennessee River system during that era.
Indeed, the apparent diversity of the Mobile Basin mollusk fauna may not so much be a consequence of evolutionary history, but rather history of a very human sort. It is best, perhaps, understood as the story of my professional hero, Calvin Goodrich (3).
 Some of my audience may recall that Calvin Goodrich began his career as a newspaperman, and that he was influenced by A. E. Ortmann to take up malacology around the years 1913 - 1917. One of Ortmann's closest professional colleagues was Herbert H. Smith, who retired in 1903 from the Carnegie Museum in Pittsburgh to become curator of the Alabama State Museum in Tuscaloosa. When H. H. Smith died in 1919, a review of Smith's collections of Alabama pleurocerids was passed by Ortmann to Goodrich.
Some of my audience may recall that Calvin Goodrich began his career as a newspaperman, and that he was influenced by A. E. Ortmann to take up malacology around the years 1913 - 1917. One of Ortmann's closest professional colleagues was Herbert H. Smith, who retired in 1903 from the Carnegie Museum in Pittsburgh to become curator of the Alabama State Museum in Tuscaloosa. When H. H. Smith died in 1919, a review of Smith's collections of Alabama pleurocerids was passed by Ortmann to Goodrich.
"The Anculosae of the Alabama River Drainage" was Calvin Goodrich's first substantial contribution to science (4). It is essentially a tribute to Smith (and indirectly to Ortmann), adding 11 new H. H. Smith nomina to a list of 14 transferred forward from Conrad, Lea, and Anthony. Goodrich described one new species himself, Anculosa smithi (named for guess who), bringing the total inventory up to 26 Leptoxis for the drainage. A concatenation of Goodrich's Plates I and II, illustrating all 26 species (5), is shown at left (and click to enlarge).
This work long predated Goodrich's (1934-41) "Studies on the Pleuroceridae," in which our hero came around to an understanding of the ecophenotypic nature of the shell morphological characters upon which pleurocerid taxonomy has historically been based. It also (of course) predated his (1939-1944) reviews of the Pleuroceridae of North America, in which Goodrich synonymized a huge fraction of the old 19th century taxonomy, combining (for example) the 26 nomina of Leptoxis (1) described from the Ohio/Tennessee basin down to nine (6). But apparently the old newspaperman could not edit his own copy. When he came back to the "Pleuroceridae of the Coosa River Basin" twenty years later (7), he preserved 18 Leptoxis species, saying, "A re-examination in 1943 has confirmed most of the decisions of 1922" (8).
By the 1940s, however, Calvin Goodrich's editorial skills were of little consequence to the Mobile Basin Pleuroceridae. Because as early as 1914 the first of seven dams was closed on the Coosa River, covering miles of the rocky rapids which were the habitat of Leptoxis with silty slackwater. And over the next 53 years, as the Coosa was almost completely impounded by Alabama Power, the Corps of Engineers was improving navigation on the Alabama River downstream with channelization, locks and dams.
Entering the 1990s, Leptoxis populations were believed to have survived in only four regions of the Mobile Basin - one tributary of the Black Warrior River (7), the upper reaches of the Cahaba River, the lower reaches of three mid-sized tributaries of the Coosa, and (amazingly) the main Alabama River downtream from the Claiborne Lock and Dam. Perhaps not surprisingly, conventional wisdom accorded snails from these four regions four different specific nomina - Leptoxis plicata, L. ampla, L. taeniata, and L. picta (respectively). The first three of these species were approved as candidate species in 1995, and after study and comment, entered the Federal lists in 1998 - L. plicata as endangered, L. taeniata and L. ampla as threatened (9). Leptoxis picta was not considered for listing.
We interrupt this extended history lesson for a brief spasm of science. I was sent large samples of all four of these nominal species (8 populations, 30 individuals per population) by our colleague Chuck Lydeard in 1996. The sample of L. picta I received was of larger, older, and more heavily-shelled animals (10), but otherwise no morphological difference between L. picta, L. ampla, and L. taeniata was apparent. My sample of Leptoxis plicata was distinctive by their higher-spired shells, bearing slight carination.
I did a proper allozyme study, comparing divergence at 9 loci among the 8 populations to 3 populations of Leptoxis praerosa, the common and well-characterized species widespread in Tennessee drainages to the north (11). The levels of genetic divergence among the picta, ampla, and taeniata populations were negligible. So it was quite clear, as of 1996, that all the Leptoxis populations known to have survived in the Alabama/Coosa River system were conspecific, L. picta being the oldest name for the group. Leptoxis plicata (of the Tombigbee/Black Warrior system) would appear to be a valid biological species.
So in summary, two of the three nominal species of Leptoxis currently on the US endangered species list are junior synonyms of a third species, which is not listed. What lessons can be learned from this mess?
Once again we see a vivid demonstration that science and public policy are two entirely different things. Most of you have heard me preach this sermon before, so I won't preach it again (12). Scientists have a language, culture, value system, and assumptions about the world that are completely different, no better or worse, from the language, culture, values, and assumptions brought by politicians, lawyers, and the natural resource managers who put laws into practice.
The adjective "endangered" is not scientific - it can't be measured, quantified, verified, or falsified. A designation of endangerment is the result of a political process, and by that process Leptoxis plicata (for example) is endangered and Leptoxis picta is not.
Now we read that yet another nominal species of Leptoxis from the Mobile Basin has been proposed for protection under the US Endangered Species Act (13). Field surveys undertaken in the upper Coosa in the late-1990s led to the discovery of a pleurocerid population in the Oostanaula River of North Georgia that has been identified as Leptoxis downei or L. foremani. In the last ten years, this population has become the target of an extensive recovery effort, our colleague Paul Johnson initiating a captive propagation program at the Tennessee Aquarium Research Institute in 2000, which he carried with him to the Alabama Aquatic Biology Center in 2005 (14).
But is Leptoxis foremani a valid biological species? Tryon synonymized foremani under L. picta in 1873, and the taxon was only resurrected by Goodrich in 1922 on the slenderest of threads (15). So given the tortured history of Leptoxis systematics in the Mobile Basin through 150 years, are we surprised that so much time, money and effort has been spent on conserving "L. foremani," while no effort whatsoever has apparently been directed toward establishing its biological reality? Nope. Science and public policy are two entirely different things.
Notes
(1) I'm going to fight the urge to digress into a discussion of the genus-level taxonomy here. Maybe one day soon. In the mean time, for the purpose of this essay, I'm lumping Anculosa and Nitocris/Mudalia together under Leptoxis.
(2) Tryon G. W. (1873) Land and Freshwater Shells of North America. Part IV, Strepomatidae. Smithsonian Miscellaneous Collections 253: 1 - 435.
(3) "The Legacy of Calvin Goodrich" See my Post of January '07.
(4) Goodrich, C. (1922) The Anculosae of the Alabama River Drainage. Misc. Publ. Univ. Mich Mus. Zool. 7: 1-57.
(5) The suviving taxa depicted on Goodrich's (1922) Plates I and II are as follows: Figs 3 - 5 are L. ampla, Figs 18-19 are L. foremani, Figs 34 - 35 are L. picta, Figs 36-38 are L. plicata, Figs 46 - 49 are L. taeniata.
(6) Goodrich, C. (1940) The Pleuroceridae of the Ohio River drainage system. Occas. Pprs. Mus. Zool. Univ. Mich 417:1 - 21.
(7) Here's another digression I hate to take. The Mobile Basin is composed of two halves - the Alabama/Coosa and the Tombigbee/Black Warrior. Most of the literature we are reviewing here is for subsets. Goodrich's (1944) "Pleuroceridae of the Coosa River Basin" would include just a geographical (not taxonomic) subset of his (1922) "Anculosae of the Alabama River," for example, and neither work would include populations like L. plicata of the Black Warrior. A map of the Mobile Basin is available [here].
(8) Goodrich, C. (1944) Pleuroceridae of the Coosa River basin. Nautilus 58: 40-8.
(9) Endangered status for three aquatic snails, and threatened status for three aquatic snails in the Mobile River Basin of Alabama. 63 FR 57610-57620 [PDF]
(10) Goodrich himself prominently noted the relationship between river size and pleurocerid shell morphology on many occasions. See my post of February '07, "Goodrichian Taxon Shift." We will return to this subject very soon.
(11) Dillon, R.T., and C. Lydeard (1998) Divergence among Mobile Basin populations of the pleurocerid snail genus, Leptoxis, estimated by allozyme electrophoresis. Malacologia 39: 111-119. [PDF]
(12) For example, "Idaho Springsnail Panel Report" (December '05), or "Red Flags, Water Resources, and Physa natricina." (March '08).
(13) Mobile Basin I: Two pleurocerids proposed for listing. Post of August '09.
(14) Interrupted Rocksnail Reintroduced to the Coosa River. Outdoor Alabama, February 2004, p. 33 [PDF]
(15) "In shell characters this species (L. foremani) is closer to A. picta Conrad than is A. formosa Lea. But while the operculum of picta and formosa are much alike, that of foremani is like the operculum of neither. The similarity of the shells of formosa and foremani, picta out of consideration, varies strangely with locality, the resemblances and differences seeming to play a game of see-saw as the collector travels down the Coosa River." (Goodrich 1922: 18).
Monday, August 24, 2009
Mobile Basin I: Two Pleurocerids Proposed for Listing
Editor's Note. This essay was subsequently published as: Dillon, R.T., Jr. (2019c) Mobile Basin I: Pleurocera puzzles. Pp 23-26 in The Freshwater Gastropods of North America Volume 3, Essays on the Prosobranchs. FWGNA Press, Charleston.
The fauna of the Mobile Basin has become a cause célèbre - perhaps the cause célèbre - of freshwater gastropod conservation in North America. I am aware of four general calls-to-arms issued in recent years that have focused on the plight of this famously diverse biota, inhabiting creeks and rivers through the length of Alabama, edging into East Mississippi and North Georgia (1). The review of Neves and his colleagues, for example, cataloged 118 freshwater gastropod species in the Mobile Basin in ten families, suggesting that as many as 38 may have been lost to extinction in the 20th century, with another 70 threatened. Yet only seven of the species remaining are currently protected under the US Endangered Species Act: four pleurocerids (Leptoxis ampla, L. plicata, L. taeniata, Goniobasis crenatella), two viviparids (Tulotoma magnifica and Lioplax cyclostomaformis) and one hydrobiid (Lepyrium showalteri). Now after a hiatus of over ten years, in late June the US Fish and Wildlife Service announced a proposal to list two new Mobile Basin pleurocerids: Leptoxis foremani and Pleurocera foremani (2). The comment period ends this Friday, August 28. Any of our colleagues on this list who might wish to offer "comments, suggestions, and any additional information on biology, threats, range, distribution, population size, or current or planned activities and the activities' possible impacts on these species or their proposed critical habitats" will find instructions on the FWS website [here].
Now after a hiatus of over ten years, in late June the US Fish and Wildlife Service announced a proposal to list two new Mobile Basin pleurocerids: Leptoxis foremani and Pleurocera foremani (2). The comment period ends this Friday, August 28. Any of our colleagues on this list who might wish to offer "comments, suggestions, and any additional information on biology, threats, range, distribution, population size, or current or planned activities and the activities' possible impacts on these species or their proposed critical habitats" will find instructions on the FWS website [here].
But what do we actually know about the freshwater gastropods of the Mobile Basin? Their taxonomy almost entirely predates the Modern Synthesis. We have seen that Calvin Goodrich, for example, seemed to have second thoughts about the 26 species of Leptoxis he catalogued in 1922 (3), later coming around to the realization that much of the shell variation expressed in North American pleurocerids may be ecophenotypic in origin (4). A bit of digging into the 6/29 issue of the Federal Register that formally proposes the new pleurocerids for listing will reveal a reference to the recent dissertation of Jeffrey Sides, reporting negligible sequence difference between P. foremani and a co-occurring species of an entirely different genus, Goniobasis ("Elimia"). What gives?
This is the first in what will probably turn out to be a sporadic series of essays on the freshwater gastropod fauna of the Mobile Basin. Between the undeniably fascinating evolutionary biology of this unique fauna and the complications of politics, personalities, and public policy there are, no doubt, many lessons to be learned.
Notes
(1) Brown, K. M & P. D. Johnson (2004) Comparative conservation ecology of pleurocerid and pulmonate gastropods of the United States. Amer. Malac. Bull. 19: 57-62. Lydeard, C. et al. (2004) The global decline of nonmarine mollusks. BioScience 54: 321 - 330. Neves, R.J., A E. Bogan, J. D. Williams, S. A. Ahlstedt, and P. W. Hartfield (1997) Status of aquatic mollusks in the southeastern United States: A downward spiral of diversity. Chapter 3 in Aquatic Fauna in Peril: the Southeastern Perspective (Benz & Collins, eds.) Southeast Aquatic Research Institute Publication 1. Lydeard, C. & R. L. Mayden (1995) A diverse and endangered aquatic ecosystem of the southeast United States. Conservation Biology 9: 800-805.
(2) Service Proposes Endangered Species Status and Critical Habitat Designations for the Georgia Pigtoe Mussel, Interrupted Rocksnail, and Rough Hornsnail: http://www.fws.gov/southeast/news/2009/r09-035.html
(3) Goodrich, C. (1922) The Anculosae of the Alabama River drainage. Misc. Publ. Mus. Zool. Univ. Mich., 7, 1-57
(4) "The legacy of Calvin Goodrich." FWGNA post of January '07. "Goodrichian Taxon Shift." FWGNA post of February '07.
The fauna of the Mobile Basin has become a cause célèbre - perhaps the cause célèbre - of freshwater gastropod conservation in North America. I am aware of four general calls-to-arms issued in recent years that have focused on the plight of this famously diverse biota, inhabiting creeks and rivers through the length of Alabama, edging into East Mississippi and North Georgia (1). The review of Neves and his colleagues, for example, cataloged 118 freshwater gastropod species in the Mobile Basin in ten families, suggesting that as many as 38 may have been lost to extinction in the 20th century, with another 70 threatened. Yet only seven of the species remaining are currently protected under the US Endangered Species Act: four pleurocerids (Leptoxis ampla, L. plicata, L. taeniata, Goniobasis crenatella), two viviparids (Tulotoma magnifica and Lioplax cyclostomaformis) and one hydrobiid (Lepyrium showalteri).
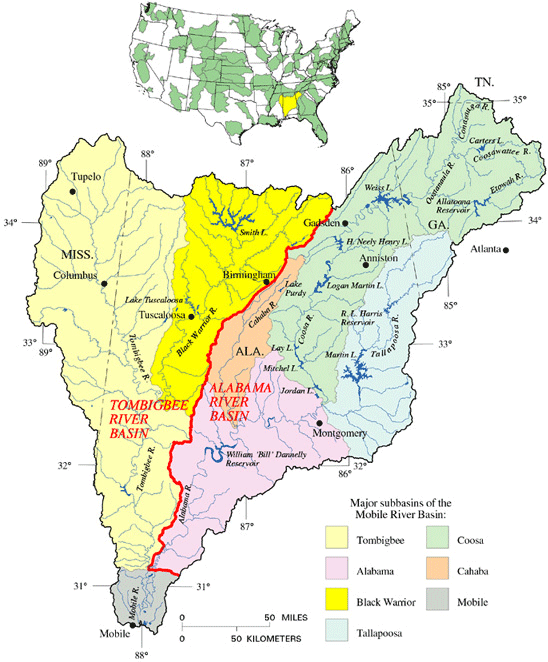 Now after a hiatus of over ten years, in late June the US Fish and Wildlife Service announced a proposal to list two new Mobile Basin pleurocerids: Leptoxis foremani and Pleurocera foremani (2). The comment period ends this Friday, August 28. Any of our colleagues on this list who might wish to offer "comments, suggestions, and any additional information on biology, threats, range, distribution, population size, or current or planned activities and the activities' possible impacts on these species or their proposed critical habitats" will find instructions on the FWS website [here].
Now after a hiatus of over ten years, in late June the US Fish and Wildlife Service announced a proposal to list two new Mobile Basin pleurocerids: Leptoxis foremani and Pleurocera foremani (2). The comment period ends this Friday, August 28. Any of our colleagues on this list who might wish to offer "comments, suggestions, and any additional information on biology, threats, range, distribution, population size, or current or planned activities and the activities' possible impacts on these species or their proposed critical habitats" will find instructions on the FWS website [here].But what do we actually know about the freshwater gastropods of the Mobile Basin? Their taxonomy almost entirely predates the Modern Synthesis. We have seen that Calvin Goodrich, for example, seemed to have second thoughts about the 26 species of Leptoxis he catalogued in 1922 (3), later coming around to the realization that much of the shell variation expressed in North American pleurocerids may be ecophenotypic in origin (4). A bit of digging into the 6/29 issue of the Federal Register that formally proposes the new pleurocerids for listing will reveal a reference to the recent dissertation of Jeffrey Sides, reporting negligible sequence difference between P. foremani and a co-occurring species of an entirely different genus, Goniobasis ("Elimia"). What gives?
This is the first in what will probably turn out to be a sporadic series of essays on the freshwater gastropod fauna of the Mobile Basin. Between the undeniably fascinating evolutionary biology of this unique fauna and the complications of politics, personalities, and public policy there are, no doubt, many lessons to be learned.
Notes
(1) Brown, K. M & P. D. Johnson (2004) Comparative conservation ecology of pleurocerid and pulmonate gastropods of the United States. Amer. Malac. Bull. 19: 57-62. Lydeard, C. et al. (2004) The global decline of nonmarine mollusks. BioScience 54: 321 - 330. Neves, R.J., A E. Bogan, J. D. Williams, S. A. Ahlstedt, and P. W. Hartfield (1997) Status of aquatic mollusks in the southeastern United States: A downward spiral of diversity. Chapter 3 in Aquatic Fauna in Peril: the Southeastern Perspective (Benz & Collins, eds.) Southeast Aquatic Research Institute Publication 1. Lydeard, C. & R. L. Mayden (1995) A diverse and endangered aquatic ecosystem of the southeast United States. Conservation Biology 9: 800-805.
(2) Service Proposes Endangered Species Status and Critical Habitat Designations for the Georgia Pigtoe Mussel, Interrupted Rocksnail, and Rough Hornsnail: http://www.fws.gov/southeast/news/2009/r09-035.html
(3) Goodrich, C. (1922) The Anculosae of the Alabama River drainage. Misc. Publ. Mus. Zool. Univ. Mich., 7, 1-57
(4) "The legacy of Calvin Goodrich." FWGNA post of January '07. "Goodrichian Taxon Shift." FWGNA post of February '07.
Subscribe to:
Posts (Atom)






{kind=link}
{kind=link}
{kind=link}
{kind=link}