Tuesday, September 22, 2015
5:57 of Physa!
I stopped by to see our good friend Bobby Martin of Martin Microscope at the Society For Freshwater Science meeting in Milwaukee this past May. And we got to talking, and one thing led to another. And two months later, out popped a six minute video of an individual Physa acuta crawling around in circles. Enjoy!
Thursday, September 3, 2015
The Lost Thesis of Samantha Flowers
Warning. The essay below is the fifth in a five-part series on species relationships in the
enigmatic North American “stagnicoline” lymnaeids. I will assume that you have read all four of
my previous posts. In addition, I also
make explicit references to two essays in my 2012 series on this same subject,
[10May12] and [4June12]. In fact, it
would probably help if you started with my [20Nov06] essay on F. C. Baker, and
read my [28Dec06] essay on the classification of the Lymnaeidae as well. Stand back, I’m going to try Science! [1]
Subsequently published as Dillon, R.T., Jr. (2019b) The lost thesis of Samantha Flowers. Pp 95-106 in Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
Insofar as I was able to determine from my vantage point
820.64 miles south of Ann Arbor, Samantha made good progress on her thesis
research through the 2012-13 academic year, and into the field season
following. I was pleased to see the
abstract of a talk she gave at the AMS meeting in The Azores in July of 2013,
although I myself was unable to attend.
And on 15Aug13 I received a very upbeat email from her, my first in over
a year. She reported that she was
“currently on the last leg of my Master's journey, prepping a manuscript to
recount my arduous tale of stagnicoline systematics that should be wrapped up
within the next month or two.” She also
promised to keep me posted “for when the sequences are thrown up on GenBank
[2].” And that was to be the last I ever
heard from her.

As that “month or two” stretched into 2014, with no reply to
my repeated emails, I began to worry that something might be amiss with our
promising young malacologist. Googling
around on the University of Michigan website, I was able to confirm that
Samantha did indeed defend her thesis, “Inferences into species delimitation of
Nearctic Stagnicola using geometric morphometric and phylogenetic methods,” on
November 15, 2013. The outcome I was
unable to determine. But surely, I thought,
if her thesis were successfully defended and signed, it must ultimately appear
for download (or purchase?) through some public outlet somewhere, yes?
No. After more than a
year of watchful waiting, in January of 2015 I finally emailed her major advisor,
Tom Duda, to inquire about the fate of Samantha and her thesis. Tom confirmed that Samantha’s 2013 defense
was indeed successful, and that he himself was surprised not to find her thesis
uploaded to the University of Michigan’s “Deep Blue” server. Apparently The University does not have firm
rules regarding the deposition of MS theses.
And Tom further confessed, “In the past we have requested that theses
and dissertations be deposited in our Mollusk Division library, but regret that
it was my oversight (in combination with her rapid departure and her not
responding to emails after she left) that got in the way of this happening with
Samantha.”
And in fact, as our conversation developed, it materialized
that Tom did not have a clean, final copy of Samantha’s thesis himself. He had apparently returned his only copy to
her with written comments, and she disappeared.
I suggested that he might check with some of the other members of her
committee, and he was able to locate a “near final form” version which he
shared with me in April. But Tom has
asked me not to distribute the document any further, since the version from
which I am working still has some errors.
Samantha’s Thesis [3] is a blockbuster. Her results
simultaneously reinforce a large and growing body of research confirming the
dramatic ecophenotypic effects of habitat on freshwater gastropod shell
morphology, and shatter 200 years of set notions about systematic relationships
in the North American stagnicolines.
Let’s digest her work in five steps.
First, Samantha’s CO1 sequence data suggest two biological
species. Perhaps some of you will recall
the review of interpopulation sequence divergence in L. stagnalis I posted on
the FWGNA blog in [4June12]. There I
argued that the general rule-of-thumb estimate of 5% CO1 divergence often
observed among biological species of pulmonate snails seems applicable to
within-continent comparisons of lymnaeid populations worldwide. This is not a law, it is a very broad-brush
guideline [4].
So below I’ve reproduced a (rather heavily-edited) version
of Samantha’s “Collapsed Bayesian-inferred CO1 tree,” with state and province
abbreviations marking samples from ME = Maine, MI = Michigan, MB = Manitoba,
and so forth. Setting aside the single
Lymnaea arctica sequence that Samantha mined from the Barcode of Life Database
[5], the ABGD prior max distance bars at right seem to suggest the two clusters
of stagnicolines I have labelled “V1” and “V2”.
Although the pairwise sequence differences between these two groups
apparently do not consistently reach 5%, an eyeball estimate from Samantha’s
scale bar, together with the plot of pairwise genetic distances Samantha
offered elsewhere in her thesis, suggests to me that they probably often do.
Note that I have modified the noun “species” with the
adjective “biological” here. This is
because populations of the two putative species seem to occur sympatically, at
least in some cases. More under my
fourth point, below.
Second, the distinction between these two putative
biological species does not coincide with taxonomic tradition, as historically
based on shell morphology. Samantha
classified each of the individual snails she sampled for her C01 analysis using
geometric morphometrics, digitizing their shell outlines with the large set of
sliding landmarks [6] shown in the colorful figure I have reproduced at the top
of this essay [7]. She recognized four
nomina by shell fatness – identifying the green and red categories as emarginata,
the gold as elodes and the blue as exilis.
The gold was an unfortunate color choice – nearly invisible between the
red and blue in her figure.

But in any case, the correct way to define any of these
nominal taxa would have been by reference to populations sampled from their
type localities. God Knows I Tried to
help Samantha with this critical component of her thesis, but for a variety of
reasons, it just didn’t work out. So I
have deleted Samantha’s specific names from the CO1 tree above [8] and
substituted simple color coding according to her morphometric analysis
[10]. And it will be obvious that the
four color categories do not correspond to the two putative biological
species. Cluster V2 shows all four
colors, and cluster V1 shows three of the four.
Third, the putative biological species do coincide with
Brady & Turner’s V1/V2 taxonomy.
Here I’ll ask you to open my essay of [10May12] in a separate window,
and refresh your memory regarding Brady & Turner’s “cryptic stagnicoline”
populations from NW Pennsylvania.
Although all four of the B&T populations inhabited fishless marshes,
and all four bore dark, skinny shells typical of elodes, their “Hartstown
Marsh” population demonstrated a consistently larger (and perhaps more
“flat-sided”) body whorl than their Conley, Osgood, and Killbuck
populations. Kip Brady’s common garden
experiments suggested that this body whorl difference seemed to be heritable.
Brady & Turner [11] considered that their Conley,
Osgood, and Killbuck populations demonstrated “typical” L. elodes shell morphology,
and called them V1. They called their
Hartstown Marsh population V2. See note
[12] for an interesting story about the example specimens figured at right below.

I forwarded samples of all four B&T populations to
Samantha in July of 2012 [13]. And sure
enough, samples from the Hartstown population (marked as PA-h on my version of
Samantha’s CO1 tree) appeared genetically distinct from the Conley, Osgood, and
Killbuck samples (marked PA-c,o and PA-k).
So although subtle, there does appear to be a shell morphological
correlate to C01 sequence divergence between the two putative species. The key character does not seem to be the
traditional fat/skinny dichotomy, but rather the relative size of the body
whorl [14].
Fourth, evidence suggests that the traditional taxonomy of
North American stagnicolines may have been based on shell characters largely
ecophenotypic in their origin. The best
example, ironically, comes from Douglas Lake, the home of the University of Michigan
Biological Field Station. Samantha
sampled 4 individuals from the waters of Douglas Lake itself, all of these
being classified as “emarginata-ovate” by her morphometric criteria, which I
have marked with green letters “d” in the C01 tree above. This small sample included three individuals
belonging to putative biological species V2, and a single individual belonging
to putative species V1. Samantha also
sampled 8 individuals from “Douglas Lake Pools,” presumably marginal ponds not
directly connected to the lake itself.
All of these individuals were classified as exilis by Samantha’s
morphometrics, and are marked with blue letters “d” above. This included 6 individuals classified as
putative species V2, and 2 classified as species V1.
Thus Samantha’s data suggest that two biological species of
stagnicoline lymnaeids seem to co-occur sympatrically in Douglas Lake, both
bearing fat shells of emarginata morphology in the main lake, and both bearing
skinny shells of exilis morphology in marginal pools. We search the world over, and sometimes the
answers we seek are right on our own doorsteps.
And fifth, we do not actually know the correct names for
either of the putative biological species.
Here I must pause, and wipe a tear from my eye. For some reason known but to God, Samantha did
not sequence that sample of topotypic L. catascopium I gave her in June of
2012. Was this tragic oversight related
to some sort of funky decision-making late in her research, regarding the
taxonomy to be employed in her thesis as a whole? See note [8] below for more.
In any case, as I have repeatedly emphasized (to Samantha,
and to you all as well!), catascopium (Say 1817) is the oldest name available
for any North American stagnicoline population.
One of Samantha’s two putative biological species almost certainly must
be catascopium by definition, and the correct name of the other species
depends.
So what to do? Almost
all of Samantha’s pale/fat snails, which might conventionally be identified as
catascopium, were classified as V2. This
set included the sample I sent her from Maine, graphed as a big green triangle
at the top of her C01 tree. And almost
all of Samantha’s V1 individuals demonstrated the dark/skinny shell morphology
conventionally associated with elodes.
So let us provisionally call the V2 species Lymnaea catascopium, leaving
the name Lymnaea elodes for putative species V1. This is admittedly a judgement call, but
seems most consistent with the taxonomy currently employed by workers in the
field.
Have I beat this horse long enough? Let me conclude with two recommendations for
further study. First, the hypotheses
advanced here can be tested with a good genetic survey of the stagnicoline
populations inhabiting the Douglas Lake area.
Somebody needs to use microsatellites, or old-fashioned allozymes, or
even older-fashioned breeding studies, to test the hypothesis that two
reproductively isolated stagnicoline species are sympatric in that lake, not
corresponding to the traditional fat catascopium / skinny elodes dichotomy, but
rather corresponding to the new V1 elodes / V2 catascopium dichotomy. And second, somebody needs to go back up the
Delaware River and fetch us some more topotypic catascopium. And find us some topotypic elodes at Lake
Canandaigua, while shopping around in Yankeeland for lymnaeids anyway. Not it.
Notes
[1] This catch phrase comes from the online comic,
xkcd.com. And although the xkcd logo
shows a stick-figure scientist flamboyantly flourishing a beaker and a calculator,
real science is at least as much theoretical as applied. To be quite precise, science is the
construction of testable models about the natural world. This essay is real science. Stand back.

[2] Not only did Samantha ultimately fail to make her MS
thesis available from any public outlet, she also failed to “throw up” any of
her sequence data on GenBank. Alas.
[3] Flowers, S. L. (2013)
Inferences into species delimitation of Nearctic Stagnicola (Gastropoda:
Lymnaeidae) using geometric morphometric and phylogenetic methods. M.Sc. Thesis, University of Michigan, Ann
Arbor.
[4] Two disclaimers.
First, gene trees are NOT species trees!
They are weak, null models of population relationship. And second, there is no cut point for
species-level sequence divergence that isn’t more exception than rule. See, for example:
- Phylogenetic Sporting and the genus Laevapex [20July07]
- Gene Trees and Species Trees [15July08]
- The Snails The Dinosaurs Saw [16Mar09]
- What is a Species Tree? [12July11]
[6] Samantha really should have digitized more than the
shell outlines. In particular, the
relative sizes of the shell whorls, especially the body whorl, seem to contain
a great deal of heritable information in freshwater gastropod populations [14],
which may be difficult to recover without landmarks on the suture lines or
aperture. See for example:
- Dunithan A, Jacquemin SJ, Pyron M (2012) Morphology of Elimia livescens (Mollusca: Pleuroceridae) in Indiana, U.S.A. covaries with environmental variation. Am Malac Bull 30:127–133.
- Dillon, R. T., S. J. Jacquemin & M. Pyron (2013) Cryptic phenotypic plasticity in populations of the freshwater prosobranch snail, Pleurocera canaliculata. Hydrobiologia 709: 117-127. [PDF]
- Dillon, R. T. & S. J. Jacquemin (2015) The heritability of shell morphometrics in the freshwater pulmonate gastropod Physa. PLoS ONE 10(4) e0121962. [PDF]
[8] Although Samantha did not explicitly cite any reference
works to support her taxonomy, it is my impression that her choices of the
category names emarginata-ovate, emarginata-narrow (“canadensis”), elodes and
exilis follow the 1992 work of Burch & Jung [9]. Whatever the origin, her
taxonomy is most unfortunate. For
unexplained (indeed unexplainable) reasons, Samantha seems to have dropped the
oldest specific nomen available for stagnicoline lymnaeids, catascopium (Say
1817), from her methods and results sections in favor of emarginata (Say
1821). But (I’m guessing here) the
decision may have come late in her project?
Because a couple samples remain identified as “catascopium” in her draft
Table 1, with purple coding in her draft Figures 10, 11, and 12. What a mess.
[9] Burch, J. B. & Jung, Y. (1992) Freshwater Snails of
the University of Michigan Biological Station Area. Walkerana 6(15): 1 – 218.
[10] It is my broad-brush impression, based on nothing more
than inspection of the figures in Samantha’s draft thesis, plus the sets of
stagnicoline shells figured by Burch and Jung, that samples our Michigan
colleagues tend to call emarginata-narrow (“canadensis”) and elodes may tend to
represent putative species V1, and the samples our colleagues call
emarginata-ovate or exilis may tend to represent V2, But since Samantha didn’t digitize the shell
aperture or suture lines, the distinction was not recovered by her
morphometrics. So let’s just focus on
Samantha’s color coding, and set her taxonomy aside to the extent possible.
[11] Brady, J. K & A. M. Turner (2010) Species-specific
effects of gastropods on leaf litter processing in pond mesocosms. Hydrobiologia 651: 93-100.
[12] For several weeks during the spring and early summer of
2012, I held cultures of the four B&T populations here in Charleston,
dissecting samples to hunt for anatomical distinctions that I ultimately did
not find. During that period both the Killbuck (V1) and Hartstown (V2)
populations laid eggs. These I hatched and
reared for quite a few months in my standard plastic aquarium boxes, at
densities that were certainly too high, largely neglecting them, changing their
water infrequently.
In any case, my results seem to confirm those of Kip
Brady. My standard culture water here in
Charleston is almost certainly much softer than that to which stagnicoline
lymnaeids are usually adapted, yielding the chalky appearances of the two
example shells figure above. Yet the V1
offspring did indeed seem to develop relatively smaller body whorls than the V2
offspring.
[13] And here’s another little confusion. Unknown to me, Kip Brady sent Jack Burch
samples from a couple of his stagnicoline populations several years prior to my
shipment to Samantha. Kip never heard
anything further. But apparently 5 of
Kip’s earlier samples were sequenced at some point along with 8 of the
individuals I sent to Samantha in 2012, and all 13 appear graphed side-by-side
in Samantha’s C01 tree, under two different labelling schemes.
[14] Body whorl differences of this subtle sort seem to be
quite heritable in freshwater pulmonates as a general rule. In fact, body whorl differences were the way
we initially distinguished Physa carolinae from Physa acuta back in 2009
[15]. For more, see:
- The Lymnaeidae 2012: A clue [9July12]
- The heritability of shell morphology in Physa h^2 = 0.819 [15Apr15]
Wednesday, August 19, 2015
The Type Locality of Lymnaea elodes
Editor’s Note. This
is the fourth installment of what I think will turn out to be a five-part
series on the tangled species relationships of the North American stagnicolines. It’s not essential, but you might find it
helpful to read my essays of [22June15], [14July15], and [29July15] before
proceeding onward.
Subsequently published as Dillon, R.T., Jr. (2019b) The type locality of Lymnaea elodes. Pp 87-93 in Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
Subsequently published as Dillon, R.T., Jr. (2019b) The type locality of Lymnaea elodes. Pp 87-93 in Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
Last month I noted that Thomas Say described two
“stagnicoline” lymnaeids in 1821, the (pale, fat) Lymnaeus emarginatus from
“lakes of Maine” and the (dark, skinny) Lymnaeus elodes [1]. Say was much more precise about the type
locality of the latter species, specifying that it “Inhabits Canandaigua
Lake.” And apparently he considered his
Lymnaeus elodes so distinct that he did not feel called to distinguish it from
any other North American freshwater gastropod [2]. The figure below was scanned from Say’s 1830 “American
Conchology” [3].
From my (perhaps selfish?) perspective almost two centuries
later, vague type localities such as that for emarginata seem preferable to
specific ones, such as that for elodes. So
much has changed here in the United States, over the last 194 years, that references
to specific type localities may well have been rendered obscure, or the
environment radically altered, and its biota extinct. But vague type localities would seem to
afford a modern researcher, such as myself, some wiggle room.

So as the vagaries of my 2012 field season played out, late
July was scheduled for an extensive survey of northern Pennsylvania, home
ported at the University of Pittsburgh’s Pymatuning Laboratory of Ecology
[4]. And I calculated that at least one
of my excursions out of PLE might take me within striking distance of the
elodes type locality on Lake Canandaigua.
And once again I had done some homework – this time, a bit
more thoroughly. Digging around in the
older literature I was pleased to unearth a charming paper written in 1932 by
my hero [5] F. C. Baker: “The Ecology of Say’s Limnaeus elodes [6].” Regarding the type locality, Baker wrote:
“It is obvious that a pond or swamp-inhabiting snail would not live in the waters of a large lake like Canandaigua and its exact ecological station must necessarily be sought in a normal habitat near the lake. No specimen of elodes has ever been found living in the lake. Many years ago the north end of the lake, where the outlet discharges its surplus waters, was an extensive swamp or marsh inhabited by elodes and other mollusks. This area has now been drained for building purposes. The southern end of the lake is also marshy. It is probable that the specimens from which Say drew up his original description came from the north end of the lake, either in the marshy tract bordering the outlet, or from a beach pool or pond bordering the upper (southern) end of the lake.”
So I arrived at the upper end of the Canandaigua Lake in the
late morning of 27July12, launched my kayak from the Woodville Access, and
paddled south to prospect for a possible “beach pool or pond bordering.” My goodness, it was swampy in there, with
luxuriant growths of floating and emergent macrophytic vegetation. I can rarely remember seeing larger
populations of Physa acuta or Lymnaea columella. I also netted a few Gyraulus deflectus and
Valvata tricarinata, both of which were interesting to my southern eyes. But I was looking for elodes habitat, which
should be isolated and essentially fishless.
And to quote my own field notes of 7/27/12 verbatim, “If there are any
vernal ponds isolated in this vast sea of cat-tails and swamp forest, one could
not find them from a kayak. The only way
to find stagnicoline habitat here would be by helicopter.”
So I paddled back to the truck and drove along the western
shore of the lake, a distance of perhaps 15 miles, noting (as Baker’s paper had
led me to expect) a “very precipitous” shoreline featuring essentially zero
habitat for freshwater gastropod populations of any sort.
And when I arrived at the lower (northern) end of the lake,
I again found it as Baker described in 1932, “drained for building purposes.” More so today, I feel sure. I did enjoy a lovely stroll in what probably
represents the vestigial remains of Baker’s “extensive swamp or marsh inhabited
by elodes,” now maintained (rather nicely) by the City of Canandaigua as
“Lagoon Park.” And I was impressed by
the aquatic vegetation in the shallow ponds protected within the park, as well
as by their large populations of bream, bluegill, and panfish of all sorts. Pleasant to the eye of the visitor,
certainly, but most inhospitable for Lymnaea elodes.

“A few years ago, while exploring the east side of the lake, a large beach pond was discovered in which were living many large pond snails of the form known as elodes. These shells are exactly like Say’s types deposited in the museum of the Philadelphia Academy and like his figures in the American Conchology… The pond (Fig 1) is situated about three miles south of the City of Canandaigua… near a group of summer cottages erected on higher ground southward.”
Baker’s Figure 1 is reproduced above. What a lovely spot, even in black and
white! Baker described the open pool as
covering about three acres, with perhaps two additional acres of “surrounding
vegetation” (which he cataloged in detail), ten “species and races” of
freshwater gastropods, and three species of pisidiid clams.
So leaving The City of Canandaigua and driving approximately
three miles south on the eastern shore of the lake, I encountered, in this
order: a “Deep Run Park,” a “Crystal Beach,” and a “Cottage City.” Deep Run Park was a nice public beach
administered by Ontario County (NY), with picnic facilities and plenty of free
parking, but no habitat for stagnicoline lymnaeids in evidence. Cottage City turned out to be a modest
community of (primarily) vacation homes dating back to the Victorian era,
perched (as Baker indicated) “on higher ground southward.” And what about that “Crystal Beach” in the
middle?
Google Earth does indeed suggest a wooded depression (or
vernal pond?) in the region of my map marked “Crystal Beach.” But alas, I could find no public access. Here is a photo I downloaded from the
Facebook page of the Crystal Beach Betterment Association:

I didn’t see any images depicting marshy or swampy areas on
the CBBA page, but comparing the location of their private property to Baker’s
(1932) description, it seems likely to me that his pond is (or was) in there somewhere
[7].
It also seems likely that with a properly-directed telephone
call or two and sufficient lead time, an interested scientist such as myself
might wrangle an invitation from the CBBA to explore their property for that
elusive topotypic population of L. elodes.
But staring at “No Trespassing” signs as the sun set on Friday evening,
27July12, this particular interested scientist was clean out of luck.
I filed my sad report with Samantha upon return to the
Pymatuning Lab that next afternoon. She
did not respond, but perhaps no response was called for. And in fact, over a year would pass before I
heard from Samantha again. I felt sure
that she was very busy up there in Ann Arbor somewhere, cranking out a forest
of gene trees, or whatever it is that the young kids do these days. Coming next time, “The Secret Thesis of
Samantha Flowers.” Stay tuned!
Notes
[1] Say, T.
(1821) Descriptions of Univalve Shells
of the United States. Journal of the Academy
of Natural Sciences 2: 149 – 179.
[2] But it is interesting to note that Thomas Say did
realize that his North American elodes was quite similar to European L.
palustris, from which he could not, apparently, distinguish it. At the bottom of his description of L.
elodes, Say noted: “This species was found by Mr. A. Jessup; it bears the most
striking resemblance to L. palustris.
The variety was found by the same enterprising mineralogist at
Morristown, New-Jersey. I have
subsequently received specimens from Mr. S. B. Collins, of New-York, who
procured them in a marsh near the Saratoga springs.” So if (as I fear) the Canandaigua elodes
population has subsequently gone extinct, I think a case could be made for
either Morristown or Saratoga Springs as a “substitute” type locality.
[3] Say, T. (1830 – 1838ish) American Conchology; or,
Descriptions of the Shells of North America. New Harmony, Indiana, “Printed at the School
Press.”
[4] A public acknowledgement of gratitude is here extended to
my good friends at The Pymatuning Laboratory of Ecology – Andy Turner, Aaron Stoler, Rick Relyea (who has since moved on) and Chris
Davis (currently in charge).
[5] For more on the life and career of this remarkable
scientist, see:
- The Legacy of Frank Collins Baker [20Nov06]
[7] For the record, here are the coordinates for my best
guess on the location of Baker’s pond: 42.8147, -77.2613. The only other nominee I can find is a
shallow pond currently between Deep Run Park and a housing development,
42.8200, -77.2586.
Wednesday, July 29, 2015
The Type Locality of Lymnaea emarginata
Editor’s Note. This post is the third installment of what I think will turn out to be a five-part series
on the tangled species relationships of the American stagnicolines. It’s not critical, but you might find it
helpful to back up and read my essays of [22June15] and [14July15] before
proceeding onward.
Subsequently published as Dillon, R.T., Jr. (2019b) The type locality of Lymnaea emarginata. Pp 81-85 in Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
Subsequently published as Dillon, R.T., Jr. (2019b) The type locality of Lymnaea emarginata. Pp 81-85 in Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
The next two North American stagnicoline lymnaeids to be
called to the attention of science, after Lymnaeus catascopium, were L.
emarginatus and L. elodes, both described by Thomas Say in 1821 [1, 2]. The figure below, clipped from Say’s “American
Conchology”[3], compares the shell morphology of emarginatus (top row) to
catascopium (bottom row). Say wrote that
emarginatus “is a somewhat larger, and considerably more ventricose species
than L. catascopium, S., and the undulation of the columella is much more
profound.” Say gave the type locality of
his emarginatus as “inhabits Lakes of Maine.”

So we left our lovely B&B in Bar Harbor on the morning
of 5July12, and by early afternoon I was collecting Lymnaea emarginata from
Pushaw Lake, about 6-8 miles north of Bangor.
The snails were quite common on rocks in about two feet of water on the
west shore of the lake. It is well that
freshwater malacology is not ordinarily so easy, lest everybody should do it.
I had actually done a bit of homework before setting off on
this little errand, and so my choice of Pushaw Lake was not entirely
arbitrary. Our good friend Scott Martin
reported emarginata populations from four counties in his (quite helpful) 1999
review, “Freshwater Snails (Mollusca: Gastropoda) of Maine [4].” So I contacted Scott for his specific
records, and among the many potential collection sites he forwarded to me was
Pushaw Lake, from which Leroy Norton reported “Stagnicola oronoensis” in
1957. Oronoensis is clearly a synonym of
emarginata.
In retrospect, however, I wish I had done more homework than
I did. A closer reading of Scott’s 1999
review would have called my attention to a 1921 paper by O. O. Nylander [5]
designating Mud Lake (in remote
Aroostook County, 150 miles north of Bangor) as the type locality for Say’s Lymnaeus emarginatus, not Pushaw
Lake. Well, as the whole project turned
out, it didn’t matter.
 |
| Rob at Pushaw Lake |
And my efforts to collect a corresponding population of
Lymnaea elodes were much more in keeping with my many years of personal
experience in the matter of the pulmonate gastropod sampling. Scott Martin actually listed six Maine
counties for dark/skinny elodes-type stagnicoline populations, but his records
were far more vague. His best locality
data were for a “Lymnaea palustris” population at the “Andrascoggin River above
Rumford.” And indeed, I found extensive,
seasonal swamps along the right bank of the river in that area, absolutely
primo habitat for mosquitos on July 5, but no sign of L. elodes. The ANSP also holds a couple collections from
the vicinity of Portland (Stroudwater and Westbrook) where I did no better.
Well, as the whole project turned out, it didn’t
matter. Oops, am I repeating myself in
my old age? This is a chord I have found
myself striking with increasing frequency, as my perspective has matured, these
latter years of my career. What we do
ain’t brain surgery, I often tell my students. It’s just snails.
So I contacted Ms. Samantha Flowers [6] immediately upon my
return to Charleston, and she was predictably pleased to hear about the bona
fide emarginata population I had collected for her from a “Lake of Maine.” She suggested that if I could ship the Pushaw
emarginata to her alive at the University of Michigan Biological Field Station
way up by Douglas Lake, she would be interested in attempting some breeding
studies with dark/skinny populations of elodes and exilis from the Michigan
area. Which (of course) I was happy to
do, out of my own pocket.
And with my July (2012) shipment to Samantha, I also
included ethanol-preserved samples of four dark/skinny stagnicoline populations
from NW Pennsylvania, which had been sent to me earlier that same the summer by
Kip Brady and Andy Turner. These were
the “cryptic stagnicolines” I first mentioned in my essay of 10May12, and
mentioned again last month. Very
mysterious, those populations…
…keep your eye on them! And tune in next month, as we continue our quest
to elucidate the tangled systematics of the enigmatic North American stagnicolines
with “The type locality of Lymnaea elodes.”
Notes
[1] Say, T.
(1821) Descriptions of Univalve Shells
of the United States. Journal of the
Academy of Natural Sciences 2: 149 – 179.
[2] To be complete, it might be argued that Thomas Say
described as many as five stagnicolines in the work cited above, including
elongatus, reflexus, and desidiosus as well as emarginata and elodes. Say realized that his nomen elongatus was
preoccupied, and changed it to umbrosus himself in his "American Conchology" [3]. The identity of desidiosus as a stagnicoline
(as opposed to a fossarine) is controversial – see the remarks of Baker (1911,
pp 318 – 321). I’m not sure what
happened to reflexa. The nomen was
passed along by Baker (1911, 1928) as perfectly valid for dark, marsh-dwelling
stagnicoline populations of very skinny shell morphology, but appears only as a
“form” of elodes in Burch (1989). I
don’t know how it got so demoted.
[3] Say, T. (1830 – 1838ish) American Conchology; or,
Descriptions of the Shells of North America.
New Harmony, Indiana, “Printed at the School Press.”
[4] Martin, S. M. (1999)
Freshwater Snails (Mollusca: Gastropoda) of Maine. Northeastern Naturalist 6: 39 – 88.
[5] Nylander, O. O. (1921)
The type localities of Lymnaea emarginata Say and L. ampla Mighels. Nautilus 34: 77-80.
[6] From this point to the conclusion of the essay, I am
assuming that you are familiar with last month’s post,
- Everything Changed When I Met Samantha [22June15]
Tuesday, July 14, 2015
The Type Locality of Lymnaea catascopium
Editor's Note. This essay was subsequently published as Dillon, R.T., Jr. (2019b) Malacological mysteries: The type locality of Lymnaea catascopium. Pp 73-80 in The Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
Although not explicitly stated, it is traditional to assume that Thomas Say was referring to his home town of Philadelphia when he wrote, in 1817, “Inhabits the Delaware River and many other waters of the United States, in considerable numbers, and may be found plentifully, during the recess of the tide, about the small streams through which the marshy grounds are drained [1].” Say was describing the habitat of the first “stagnicoline” lymnaeid, Lymnaea catascopium. Standing on the Philadelphia waterfront in June of 2012, however, I found it nearly impossible to imagine how any self-respecting freshwater gastropod population of any description might ever have inhabited such a place.
 |
| Philadelphia 1702, from phillywatersheds.org |
The Delaware is reliably fresh but quite tidal at
Philadelphia, with a daily range of several feet. So I gather that in the 18th and
early 19th centuries, merchant ships anchored some distance offshore
and transmitted their cargos over these "marshy grounds" at high tide by tender. Then as the years advanced and the technology
improved, the river must have been dredged and the fill material dumped
directly onshore, creating more land, deepening the harbor, and allowing direct offloading of ships to finger-like cargo docks. Whatever the historical scenario, however, by
June of 2012 it was clear to this particular 21st-century
malacologist that his efforts to sample a topotypic population of L.
catascopium from the Delaware River must be re-directed upstream.
 |
| Philadelphia 1908, from phillywatersheds.org |
The Academy of Natural Sciences holds one lot of
L. catascopium collected by Charlie Wurtz from Pennypack Creek in 1948. Pennypack Creek drains an entirely urban
(actually, rather post-industrial) catchment inside the Philadelphia city
limits, emptying into the Delaware just a few miles upstream from the
docks. And so it was to Pennypack Park that
I set my GPS early in the morning of June 11, 2012.
I threw my kayak directly into the Delaware River
and paddled upstream through the mouth of the creek into a zone of broad,
intertidal mudflats decorated with a dense stand of arrowhead (Sagittaria). The creek narrowed and deepened substantially
as I paddled upstream, looking for the solid substrates I knew that populations
of Lymnaea catascopium require. Soon the
air crackled with gunfire, as I passed alongside (perhaps “beneath” would be a
better preposition) the Philadelphia Police Academy Firing Range. After about a mile the creek had shallowed to
the point I could get out and walk.
But I found no L. catascopium, nor indeed any
habitat. The steam bed was too
muddy. I found Littoridinops moderately
common on floating debris, a few Physa acuta, N=1 Amnicola, and some beer-can limpets, but that was it. So I paddled
back downstream to the truck, loaded my kayak, and drove a couple miles
upstream to the Verree Road Bridge.
Pennypack Creek was lovely at that point on a June afternoon, but flashy
and low-nutrient, and simply not the kind of place one might expect L. catascopium.
Some nontrivial fraction of the early 19th
century prosperity of Philadelphia was due to the network of canals
communicating between the Delaware River and the interior of rapidly-expanding
America. In 1832 the Delaware Canal was
completed to run 60 miles along the right (descending) bank of the river from
Easton to the quaint old town of Bristol, PA, about 15 - 20 miles above
Philadelphia. And the Delaware and Raritan
Canal was completed in 1834, connecting New Brunswick, NJ, to Bordentown, and climbing the left (descending) bank of the Delaware. So on 12 June I checked the
historic lock areas around Bristol, and also across the river at D & R
Canal Lock #1, at Bordentown. The latter spot was tough to access
but afforded a pretty and diverse habitat with a disappointingly poor
freshwater gastropod fauna. The still-rather-strongly
tidal environment is probably a factor.
Just a couple Physa acuta, and a Menetus or two, and I was gone.

The Delaware River passes through the fall zone at
Washington Crossing, PA, famous primarily as the type locality of Physa
ancillaria (Say, 1825). Breeding results
we published back in 2006 suggested that ancillaria is a fattish shell morph of
Physa gyrina [2]. Most interestingly,
some allozyme gels we ran in support of that research effort in 2005 returned
evidence of low-frequency hybridization between P. gyrina and P. acuta at
Washington Crossing – the only place (to this day) where the phenomenon has
been documented. You would think that at
least a couple of the numerous historical markers one finds on both sides of
the Delaware River at this point would feature such a remarkable finding. But no.
Both Physa gyrina and P. acuta are common in the rocky pools
at Washington Crossing, as they are upstream for several hundred miles. The Delaware River is one of the few places
where the two species are so richly sympatric, in my experience. Also making an initial (or possibly final?)
appearance at the fall line is Pleurocera virginica. But no evidence of Lymnaea catascopium
whatsoever.
The gastropod fauna continued to richen as I
collected my way north upstream the next day.
The list lengthened to include Helisoma trivolvis, H. anceps, Gyraulus,
Laevapex, both species of Ferrissia, Lyogyrus, and even (ultimately, way up
north) Somatogyrus. The most memorable
snapshot from my June 13 field experience was the aquaviaduct at Tohickon Creek
– a genuine marvel of 19th century engineering. Here barges plying the Delaware Canal would
have passed through a covered bridge (or trough, maybe a better noun)
perpendicular to and 20 feet above the rocky creek below. I passed through the Delaware Water Gap as
the sun set on the third day of my efforts to collect L. catascopium from its
type locality.
 |
| Tohickon Aqua-viaduct (PA DCNR) |
I really needed a sample of at least 30 individuals
to estimate allozyme frequencies. So I
redoubled my efforts in all similar habitats and substrates around Milford
Beach over a period of about an hour.
But alas, no additional specimens came to light. So I drove 25 miles upstream to Shohola
Bridge, where two hours’ effort netted an additional N=5 juvenile catascopium. Further upstream at Metamoras Boat Landing
and Dillontown the river did not seem as rich, and I struck out.
 |
| Delaware Water Gap |
If my efforts to collect a decent topotypic sample
of L. catascopium were less than successful, however, my parallel efforts to
find a matching population of L. elodes were an abject failure. I had several leads. In fact, the naturalist at Echo Hill
Environmental Education Center near Lebanon, NJ, had sent me a sample of L. elodes
for identification in December of 2010.
But I stomped all around in the wooded swamp where the specimens were
collected, and couldn’t find so much as a shell.
I confess that I may not have been in the best of spirits
at the AMS welcome mixer Sunday evening, when up walked our good friend Tom
Duda, with a nice young lady in tow. Tom
introduced her as Ms. Samantha Flowers, a new graduate student at the
University of Michigan [3]. And Samantha
had chosen as her research project – if you can believe this – the evolutionary
relationships among the stagnicoline lymnaeids.
 |
| Have you seen me? |
She related to me, as the conversation unfolded, that
she planned to use a variety of approaches, including molecular phylogenetics
and geometric morphometrics, and sample as broad a range of catascopium, emarginata,
elodes, and exilis populations as time and resources permitted. I’m not crazy about gene trees, I thought to
myself [4], but they do work with small sample sizes. And what can I myself do with my crappy
little sample of N=7 topotypic Lymnaea catescopium except go back up the
Delaware again and try to find 23 more?
And such a nice young lady! So bright, and so eager to learn! In five minutes not only had I decided to
give her my sample of topotypic L. catascopium, I had resolved to help her with
the rest of her thesis in any way I could.
The next morning I transmitted my little sample of
L. catascopium to Samantha, and told her she could keep my half-gallon thermos
jug to carry them home in. I also
promised to her that I would continue to move forward on my original study
design, and that I would try to send her additional samples as the summer
progressed. Looking back on it, I wasn’t
entirely sure that she appreciated the potential for ecophenotypic plasticity
in her chosen study organisms, or indeed that she actually understood the
design of the study I (we?) were working on.
In fact, I was not entirely sure she understood
the significance of the little sample of snails I handed her that morning. Thomas Say’s (1817) nomen “Lymnaea
catascopium” is the oldest available name for any of the North American
stagnicolines. Which means that
regardless of all the other names invented by all the other malacologists to
name all the other stagnicoline populations in all the other regions of the
United States and Canada, any population matching those N=7 crappy little snails
in that red jug must be Lymnaea catascopium by definition. They were her control. Every other sample she might acquire would be
an experiment.
With the benefit of three years’ hindsight, I
think that it was probably too early in Samantha’s professional career for her
to take this all in. But stay tuned! Coming up next month - the type localities of
L. elodes and L. emarginata.
Notes
[1] Lymnaea catascopium was one of a long list of species
that Thomas Say described in the entry entitled, “Conchology,” which he
contributed to Nicholson’s British Encyclopedia of Arts and Sciences. Nicholson’s Encyclopedia was published at
Philadelphia in three editions: 1816, 1818, and 1819. I gather that these works are very rare in libraries today. And I also gather that the "1816 Edition" was actually published in 1817. I myself only have access to W.
G. Binny’s (1858) secondary reference entitled, “Complete Writings of Thomas Say on
the Conchology of the United States.”
And Binny only reprints the third (1819) edition. So that’s where I got the quote above.
[2] Dillon, R. T., and A. R. Wethington.
(2006) No-choice mating experiments
among six nominal taxa of the subgenus Physella (Basommatophora:
Physidae). Heldia 6: 41 - 50. [PDF]
[3] From this point onward in the present essay, I
am assuming that you have read last month’s post,
- Everything Changed When I Met Samantha [22June15]
[4] This is a long-running theme on the FWGNA blog, for example:
- Gene Trees and Species Trees [15July08]
Monday, June 22, 2015
Everything Changed When I Met Samantha
Editor's Note. This essay was subsequently published as Dillon, R.T., Jr. (2019b) Everything changed when I met Samantha. Pp 67-72 in The Freshwater Gastropods of North America Volume 2, Essays on the Pulmonates. FWGNA Press, Charleston.
Three years ago I posted a series of five essays on this blog entitled, “The Lymnaeidae 2012” [1-5]. My primary motivation was the imminent expansion of the FWGNA project into the Mid-Atlantic States, where I expected that we would come into contact with the range of an enigmatic group of lymnaeids called the “stagnicolines.” And a couple comprehensive molecular phylogenetic studies had also recently been published that I thought might cast some light on systematic relationships in the group.
Three years ago I posted a series of five essays on this blog entitled, “The Lymnaeidae 2012” [1-5]. My primary motivation was the imminent expansion of the FWGNA project into the Mid-Atlantic States, where I expected that we would come into contact with the range of an enigmatic group of lymnaeids called the “stagnicolines.” And a couple comprehensive molecular phylogenetic studies had also recently been published that I thought might cast some light on systematic relationships in the group.
 |
| Samantha |
Who was Ms. Samantha Flowers? And where has she gone, long time
passing? Our good friend Tom Duda from
the University of Michigan introduced her at the AMS meeting in Cherry Hill in
June of 2012, and we kept in touch until August of 2013, at which point she
disappeared. But she left behind a
tangled body of potentially important research on the genetics of the stagnicolines,
which we will sort through together, as this, our fresh series of essays on the
Lymnaeidae unfolds.
So when last we left our story, it may be recalled
that the Baker/Burch system for the classification of the North American
Lymnaeidae recognizes 21 species of stagnicoline lymnaeids in two subgroups. The dark-bodied populations of bogs, marshes
and vernal ponds bearing slender shells include elodes (Say 1821), exilis (Lea
1834) and three others more recently described.
The pale-bodied inhabitants of open waters, bearing broader, more robust
shells include catascopium (Say 1816), emarginata (Say 1821) and 14 others more
recently described.
Half of the challenge with which we wrestled in
2012 was the relationship between our New World stagnicolines and those of the
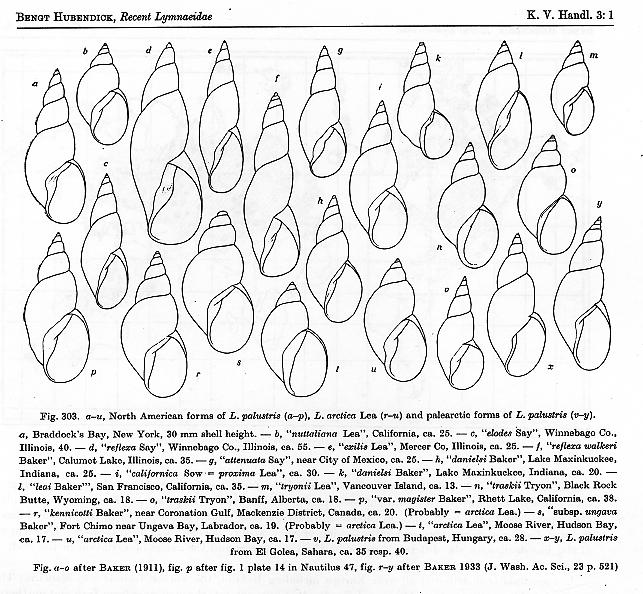
Old. In 1951, my hero Bengt Hubendick
synonymized all the dark/skinny species of North America under the European
palustris (Muller 1774). Hubendick’s
figure of L. palustris is reproduced below – click for a full-sized version,
with caption [6]. But by the 1960s evidence
had begun to accumulate that the European palustris is a complex of several
cryptic species, distinguishable only by detail of reproductive anatomy. And in my essay of 10May12 [2], I offered
evidence that at least two cryptic species of dark/skinny stagnicolines also seem
to inhabit the ephemeral ponds and marshes of NW Pennsylvania.
Although I did not mention it at the time, shortly
after I published my “cryptic stagnicoline” essay of 10May12, I asked my good
friends Kip Brady and Andy Turner to send samples of their enigmatic Pennsylvania
populations to Charleston, which I dissected, comparing details of their reproductive
anatomy to figures from the European literature. Alas, I was unable to distinguish any of
these populations anatomically, and let the matter drop. But the cryptic stagnicolines of Brady &
Turner turned out to be key to disentangling Samantha Flowers’ research
results, when we were finally able to examine them in 2015. So keep this in the back of your mind.
The other half of the challenge to working out the
systematic relationships among our American stagnicolines is their great potential
for ecophenotypic plasticity of shell.
Given the large body of research results such as those of Christer
Bronmark on European Lymnaea peregra (aka “Radix balthica”), it is not
inconceivable that the robust shells with enlarged body whorls born by
populations we call catascopium or emarginata here in North America arise as an
ecophenotypic response to life on solid substrates in open waters, exposed to
fish predation [4]. There may be no
additively heritable basis for the distinction between the broad, heavy shells
of the catascopium/emarginata subgroup and the slender, gracile shells of the elodes/exilis
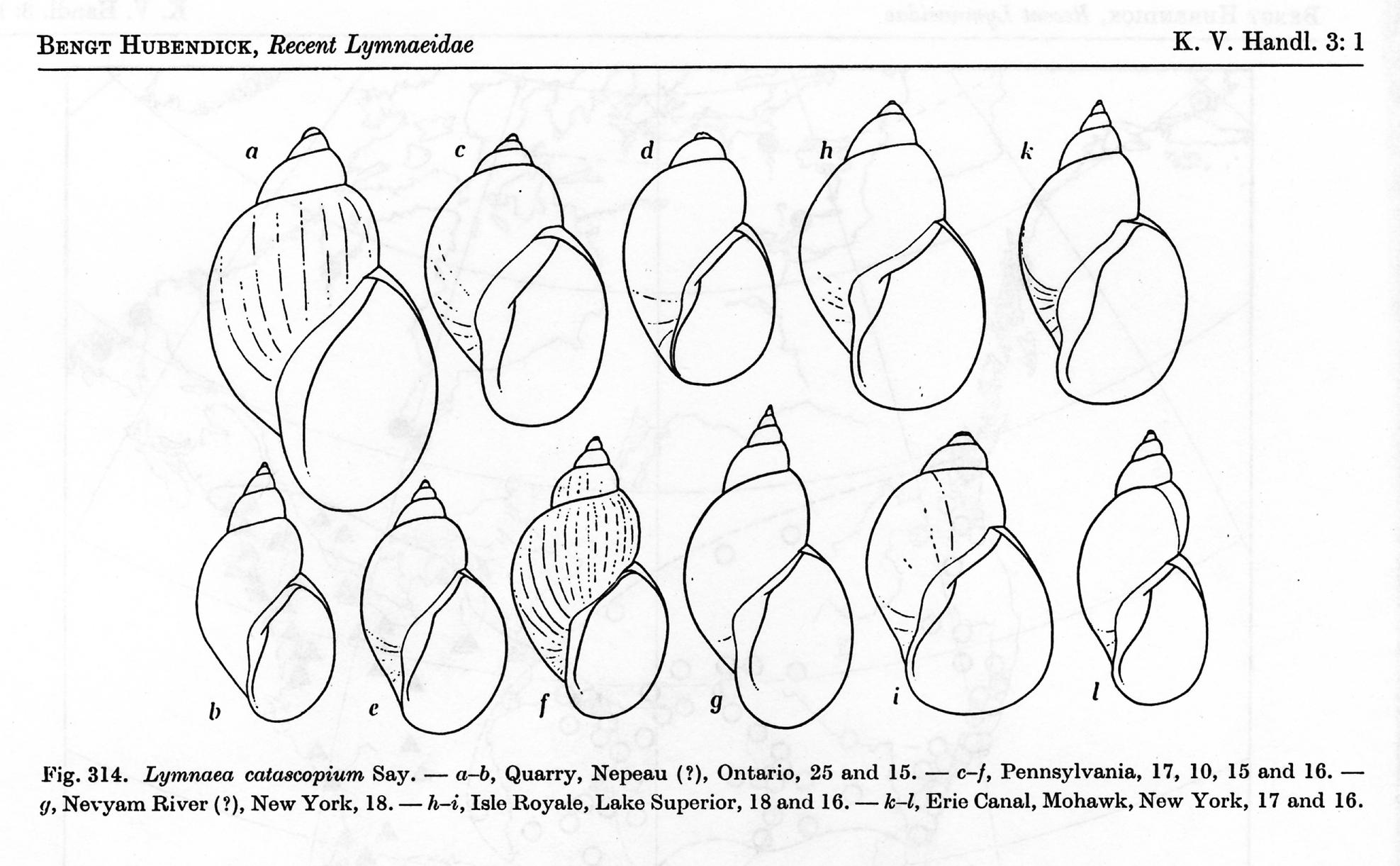
subgroup whatsoever. Hubendick’s figure
of L. catascopium is reproduced below – click for a full sized version, with
caption.
So with the potential for cryptic speciation and shell
ecophenotypic plasticity firmly in mind, in the spring of 2012 I designed a genetic
survey of the North American stagnicolines.
My plan was to sample populations from the type localities of the four
oldest nomina – catascopium, emarginata, elodes, and reflexa. And along with each topotypic population, I
also hoped to sample a nearby population bearing the opposite shell form. My hypothesis was that each broad, heavy
open-water population would prove most genetically similar to its local
slender/gracile marsh-dwelling population.
For example, the type locality of L. catascopium
is the Delaware River at Philadelphia, and the type locality of L. emarginata
is “Lakes of Maine.” In 2012 it seemed
likely to me that Delaware River catascopium might prove most genetically
similar to the populations of (nominal) L. elodes that I expected to find in
the marshes of Delaware tributaries in eastern Pennsylvania, and that Maine emarginata
might prove most genetically similar to populations of nominal L. elodes
sampled from the marshes and vernal ponds of Maine. I imagine my readership will recognize this
study design as the same I have used to confirm “cryptic phenotypic plasticity”
in a variety of pleurocerid taxa in recent years [7].
And so I mapped out an itinerary for my 2012 field
season. I set aside six days in June for what (I presciently imagined)
might be a challenging quest to re-discover L. catascopium in the Delaware
River, and L. elodes in vernal habitats of the Delaware Valley, after which I
planned to attend the meeting of the American Malacological Society,
conveniently scheduled in the Philadelphia suburb of Cherry Hill (NJ) June 16 –
21.
Everything changed when I met Samantha. But coming next month… “The type locality of
Lymnaea catascopium.”
Notes
[5] The Lymnaeidae 2012: Fossarine Football [7Aug12]
[6] Hubendick, B. (1951) Recent Lymnaeidae, Their Variation, morphology, taxonomy, nomenclature and distribution. Kungl. Svenska Vetenskapskademiens Handlingar. Fjarde Serien Band 3, No. 1. Stockholm: Almquist & Wiksells.
See The Classification of the Lymnaeidae [28Dec06]
[6] Hubendick, B. (1951) Recent Lymnaeidae, Their Variation, morphology, taxonomy, nomenclature and distribution. Kungl. Svenska Vetenskapskademiens Handlingar. Fjarde Serien Band 3, No. 1. Stockholm: Almquist & Wiksells.
See The Classification of the Lymnaeidae [28Dec06]
Monday, May 11, 2015
Cornhusker Freshwater Gastropods
Editor’s Note. This essay was subsequently published as:
Dillon, R.T., Jr. (2019d) Cornhusker Freshwater Gastropods. Pp 229 - 233 in The Freshwater
Gastropods of North America Volume 4, Essays on Ecology and Biogeography. FWGNA Press, Charleston.
Kudos to our good friend Bruce Stephen for his thorough
review of the freshwater gastropod fauna of Nebraska, just published in the
most recent issue of the American Malacological Bulletin. A pdf download is available from the link
below [1].
The surface waters of Nebraska drain entirely to the east, primarily
through the broad, sandy Platte River, as well as through the Niobara and
Republican Rivers, all tracing slow, braided paths toward the Missouri. Glaciers extended over the greener, eastern
quarter of the state about 10,000 years ago, although the western half of the
state seems to have remained a bit too dry.
Today about 40% of the state is planted in row crops, dependent largely
on the miracle of modern irrigation. The
other 60% of Nebraska is sand hills and semi-arid plains, typically given over
to ranching.

I am sure that there are many nice things that could be
said about Nebraska and its fine citizenry, but the state does not strike me as
especially rich in habitat for freshwater gastropods. So I was mildly surprised to see that our
buddy Bruce’s list extended to N = 31 species, from three primary (and hoary!) sources:
Tryon 1866, Aughey 1877, and Walker 1906, a scattering of more recent
references, and the collections at the University of Nebraska Sate Museum [2].
Actually, the starting number of species exceeded 80. The biggest challenges faced by our good
buddy were the pruning of over 40 synonyms from the 19th-century
taxonomy, and the weeding of around 8 - 10 valid nomina apparently introduced
to the fields by error. The taxonomy of
the 31 species that emerged from Bruce’s intensive efforts with hoe and hook was
neat, clean, and modern [3].
Loyal followers of this blog may remember my post of 20Apr06,
entitled “Surveying the Heartland” [4].
That essay was prompted by our good friend Tim Stewart’s review of the
freshwater gastropods of Iowa, very similar in scope and methodology to the
present work by Bruce Stephen. And in my
post of 23Jan09, I compared the (46 species) fauna of Iowa to that of Indiana in the east and Missouri in the south, identifying 20 species
shared by the three boxy Midwestern states, 15 uniquely shared with the former
and 6 uniquely shared with the latter [4].
The FWGNA readership may also remember my post of 26Nov08
reviewing a popular guidebook on the freshwater gastropods of Colorado, based
on the (1989) work of Shi-Kuei Wu [4]. So
with the publication of the present review of the Nebraska fauna, we have
available an unbroken transect, extending 1,000 miles from the banks of the
Mississippi River at Davenport, across the Great Plains and over the Rocky
Mountains to the Colorado River at Grand Junction.
 Should we expect to discover that any of our buddy Bruce’s
31 species are unique to Nebraska? Or
might we find the Nebraska fauna perfectly transitional between those of Iowa
and Colorado? The figure at left is a Venn
diagram of boxy western states, composed using the same convention I pioneered
in my essay of 23Jan09. The areas of the
states are adjusted relative to their freshwater gastropod faunas – the 44 species
documented from Iowa [5] rendering it over twice the size of Colorado (with 20
species, ref. 6), Nebraska in the middle.
Should we expect to discover that any of our buddy Bruce’s
31 species are unique to Nebraska? Or
might we find the Nebraska fauna perfectly transitional between those of Iowa
and Colorado? The figure at left is a Venn
diagram of boxy western states, composed using the same convention I pioneered
in my essay of 23Jan09. The areas of the
states are adjusted relative to their freshwater gastropod faunas – the 44 species
documented from Iowa [5] rendering it over twice the size of Colorado (with 20
species, ref. 6), Nebraska in the middle.
And we do indeed see a rather smooth malacological
transition (perhaps “attenuation” would be more descriptive) moving west from the relatively
rich fauna of Iowa to rather poor Colorado.
Setting aside the 16 cosmopolitan species shared by all three states,
Nebraska shares 12 uniquely with Iowa and just one with Colorado (Lymnaea
bulimoides). But again perhaps
surprisingly, Nebraska does seem to boast two freshwater gastropod species found in
neither Iowa nor Colorado: Physa pomilia and Gyraulus crista.
Our buddy Bruce was a bit dubious about the Physa pomilia
records, as am I. The 15 records from S-K Wu
(2004-05) predate our modern understanding of the taxon [7]. But the single G. crista record (from Taylor
1960) appears to be bona fide, this (rather boreal) species being documented
from Wyoming to the northwest.
Science is the construction of testable models about the
natural world. This definition I am in
the habit of repeating once a week to my Genetics Lab 305L students, 14 weeks
per semester. Historical reviews such as the work just published by our buddy Bruce Stephens serve the important function of providing models,
which we malacologists of the present day really ought to test. The job most certainly is not over in
Nebraska, or in Iowa, or indeed in most of the United States of America, sea to
shining sea. But it is well begun.
Notes
[1] Stephen, B. J. (2015)
Species composition of Nebraska’s freshwater gastropod fauna: A review
of historical records. American
Malacological Bulletin 33: 61 – 71. [PDF]
[2] The author’s online queries of the collection
databases at the USNM, ANSP, UMMZ, and several museums of more regional
character field to yield a single freshwater gastropod record from Nebraska.
[3] Well, there are a couple smudges here and there, but I won’t
quibble.
[4] My previous posts relevant to the biogeography of Western
and Midwestern states, in order of appearance, have been:
- Surveying the Heartland [20Apr06]
- Review: Field Guide to the Freshwater Mollusks of Colorado [26Nov08]
- The Freshwater Gastropods of Indiana [23Jan09]
[6] Wu’s (1989) monograph actually listed 30 freshwater
gastropod species. I have collapsed 15
of his specific nomina into five names as follows. Lymnaea humilis = obrussa + parva. Physa acuta = anatina + cupreonitans +
gyrina. Physa gyrina = elliptica +
heterostropha + integra + utahensis.
Helisoma trivolvis = trivolvis + scalare + subcrenatum. Ferrissia fragilis = fragilis + walkeri. For more on the physid situation in Colorado,
see my essay of [14Oct08].
[7] Dillon, R. T.,
J. D. Robinson, and A. R. Wethington (2007)
Empirical estimates of reproductive isolation among the freshwater
pulmonates Physa acuta, P. pomilia, and P. hendersoni. Malacologia 49: 283 - 292. [PDF]
Subscribe to:
Posts (Atom)





