Editor’s Notes - If you are looking for something citable, the gray-literature report upon which the following essay is based can be downloaded as FWGNA Circular #6 from footnote [1] below. This essay was subsequently published as: Dillon, R.T., Jr. (2023b) Just 125 species of Pyrgulopsis in the American West. Pp 233 – 242 in The Freshwater Gastropods of North America Volume 6, Yankees at The Gap, and Other Essays. FWGNA Project, Charleston, SC.
In July [6July22] we reviewed the career of the USNM-Smithsonian’s Dr. Robert Hershler, for over 30 years the undisputed authority on the hydrobioid freshwater gastropods of North America. Running our finger through the checklist that my buddy Bob published with Hsiu-Ping Liu in 2017 [2], we counted “126 species of Pyrgulopsis inhabiting the waters of the Western United States, 107 of which Bob Hershler was the author.”
Journey back with me now to a bleak Tuesday morning in May of 2020. The ivory towers of Academia were locked and bolted, the store shelves stripped bare of toilet paper [3], the streets silent save for the rattle of wooden handcarts, and rhythmic appeals to bring out the dead. And I opened my email inbox and found a message from Hsiu-Ping. She asked me if I would be “interested in working on a project to determine the species status of Pyrgulopsis vinyardi and P. gibba.” And here was my reply:
“Well, OK, maybe. It would take me a while to get up to speed on the Western hydrobioids, and the maximum speed I could ever achieve would look like standing still, next to Bob Hershler. But Bob’s not taking any more laps around the track, and I am.”
Thus encouraged, sort-of, Hsiu-Ping proceeded to lay out the situation. Bob had described Pyrgulopsis gibba from the northwestern Great Basin Desert of California, Nevada, and Oregon in a relatively small paper published in 1995 [4] and followed with a description of P. vinyardi endemic to "two springs in the Squaw Valley drainage" of north-central Nevada in his big monograph of 1998 [5]. The two bore shells strikingly different in the relative size of their body whorls, and penises notably different in their morphologies as well.
 |
| Pyrgulopsis vinyardi [5] and P. gibba [4] |
Regarding penial morphology, see the figure below. I have marked the part of the penis that actually does the job, really just a simple filament, with the red letter “P.” Everything else in Bob’s dorsal-and-ventral figures labelled P. vinyardi (boxed), and in his three dorsal-and-ventral figures labelled P. gibba, is the ridiculously enlarged and elaborate penial lobe characteristic of Pyrgulopsis. If you’re curious to see a whole mount of the actual organ itself, look back at my July post [6July22].
The distribution of glandular regions on the surface of this spatulate or blade-shaped lobe has for many years been considered diagnostic of hydrobiid species. Bob has marked dorsal glands as “Dg,” ventral glands as “Vg” and terminal glands as “Tg.”
Bob wrote in his description of P. gibba: “This species is unique among members of the genus (as of 1995) in having penial ornament of terminal gland, Dg3, and ventral gland.” He went on to observe, however, “Dg3 often present, either as a small papule (sometimes double) or large raised unit.” Note the modifier, “often.” Bob figured one P. gibba penis that had no Dg3 at all. See the Dg3 regions encircled in red. And in 1998 he noted that the development of the ventral gland often varies as well.
 |
| Penial morphology, modified from Hershler [6] |
The obvious analogy is to “lock-and-key” reproductive isolation such as has been widely documented across the Phylum Arthropoda, except that the gastropod lock is a bag, and the gastropod key is a sock.
 |
| I'm not buyin' it! |
And besides. Anybody who has walked through the weeds by a pond on a warm summer night, or hell, anybody who has a friend with a male dachshund, you all know. Males will do it with anything. Coke bottles. It just does not matter.
So, both Bob’s 1995 description of P. gibba and his 1998 description of P. vinyardi were published absent any genetic data, before he met Hsiu-Ping. Between 2003 and 2008 Bob and Hsiu-Ping did, however, publish mitochondrial CO1 gene sequences for three individual P. gibba and one P. vinyardi [9]. And it materialized that gibba and vinyardi are very similar genetically, mtDNA percent sequence divergence ranging just 0.5% to 1.1%.
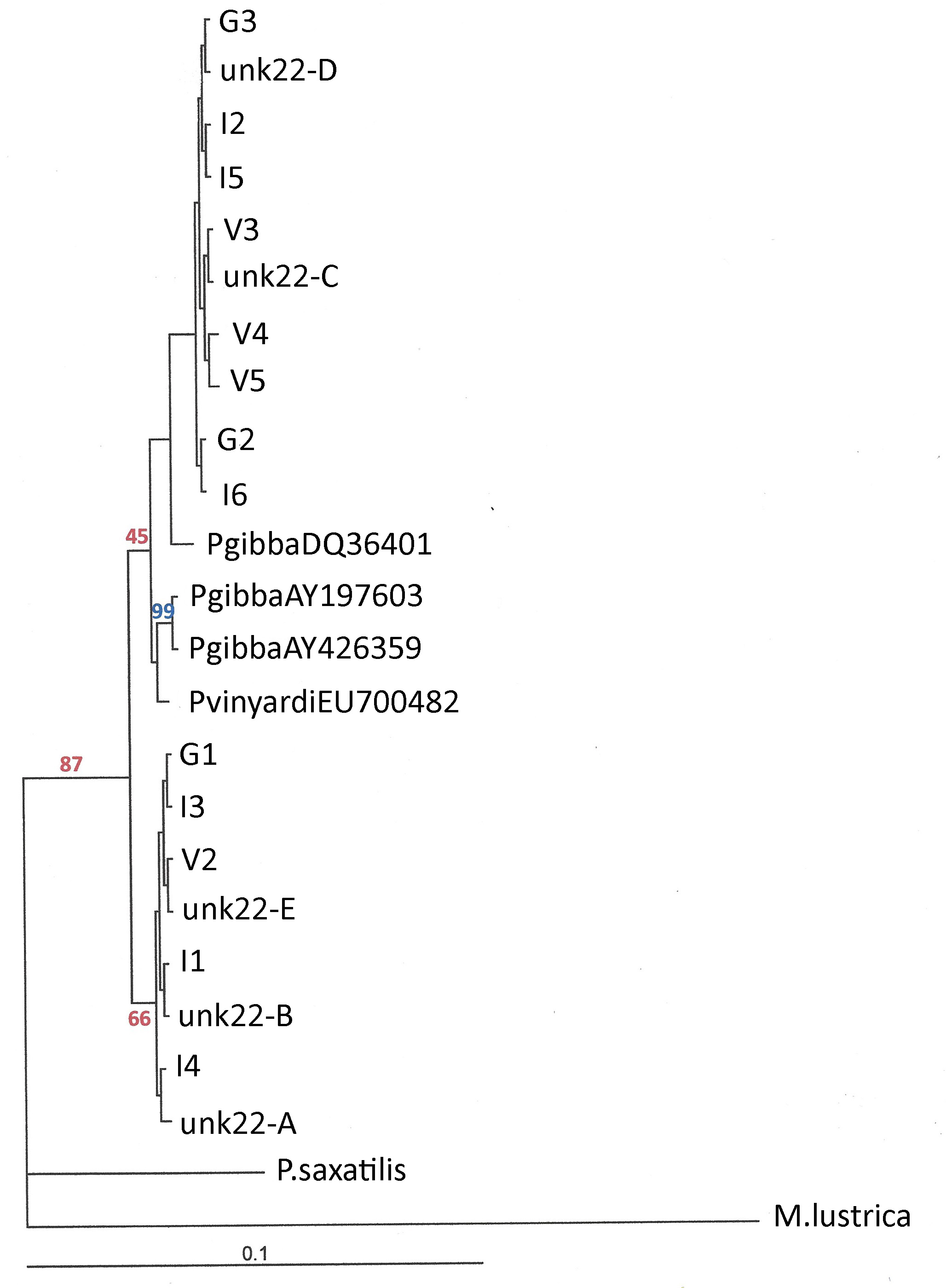
All of which now brings us back up to the dark days of May 2020, and my email exchange with Hsiu-Ping. Hsiu-Ping explained to me that she had recently agreed to provide molecular identifications for a set of 21 Pyrgulopsis samples [10] collected from northern Nevada by Ms. Diana Eck of the environmental consulting firm, Stantec. And that she had sequenced the CO1 gene from approximately 4 – 6 individuals from each population, for a total sample size of N = 88. The Baysian tree below shows the N = 29 unique CO1 haplotypes Hsiu-Ping discovered, with unidentified population number (“unk”), setting aside duplicates. Also shown are the three control P. gibba sequences from GenBank, and the one control P. vinyardi.
 |
| CO1 sequence diversity in N. Nevada Pyrgulopsis |
So, the second branch of the tree does indeed divide the control vinyardi from the control gibba, as one might expect. But look at the samples from unidentified population #22, collected 23 miles NE of Lovelock, Nevada. Sequences 22-A and 22-B cluster with vinyardi, while sequence 22-C clusters with gibba! Could this be evidence that Spring #22 is inhabited by both P. gibba and P. vinyardi? And that the two populations demonstrate reproductive isolation in sympatry? My knowledge of the vast and weighty literature is far from encyclopedic, but I cannot recall any case of sympatric Pyrgulopsis species ever previously documented in the American West.
Hsiu-Ping and I resolved to test population #22 for character phase disequilibrium [11]. And so it came to pass that in June of 2020 a fresh sample of Pyrgulopsis collected from population #22 arrived on my doorstep, courtesy of Ms. Eck. My half of the study was to dissect these snails and characterize their penial morphology as either matching P. vinyardi or matching P. gibba. Then sending the residual tissues to Hsiu-Ping, she would characterize the individuals as either matching vinyardi or matching gibba by their CO1 gene sequence. A significant relationship between penial morphology and CO1 sequence would suggest reproductive isolation within the sample, confirming the specific distinction between vinyardi and gibba.
 |
| Pyrgulopsis from Site 22 |
And so, I went to work with tiny forceps and even tinier dissecting needles, cracking and dissecting 30 adults [12], identifying 15 females and 15 males. I ignored dg3, which is an unreliable character. Then five males demonstrated both dg1 and dg2, matching P. vinyardi. Three males did not demonstrate either dg1 or dg2, matching P. gibba. And seven males demonstrated either dg1 or dg2, intermediate between gibba and vinyardi. And in July of 2020 I forwarded 15 little tubes onward to Hsiu-Ping, 5 marked G for gibba, 3 marked V for vinyardi, and 7 marked I for intermediate.
The Baysian tree below shows Hsiu-Ping’s results for 13 of the 15 snails I dissected (setting aside 2 duplicate sequences), plus all five of the sequences she obtained from Ms. Eck’s original sample, plus the four control sequences from GenBank. Hsiu-Ping’s analysis did resolve two sort-of distinct clusters of CO1 sequence within population #22, but those two clusters did not correspond to penial morphology, nor indeed, did they correspond especially well to the four CO1 sequences previously deposited in GenBank, three from nominal gibba and one from nominal vinyardi. There is no pattern in the distribution of samples labelled G, V, and I.
Hence there is no evidence of character-phase disequilibrium between penial morphology and CO1 sequence in Pyrgulopsis population #22. Hence there is no evidence of reproductive isolation between P. gibba and P. vinyardi. The gray-literature report we filed with Ms. Eck on 22Oct21, available for download as FWGNA Circular #6 from footnote [1] below, concluded “that P. gibba and P. vinyardi should be synonymized into one species.” My buddy Bob’s (1995) gibba would have priority over his (1998) vinyardi.
 |
| From Liu & Dillon [1] |
OK, I know that’s a lot of technical detail for a silly, frivolous blog post. So come back up to the surface with me and let’s take a big, fresh breath of air together. Look at those two little shells I figured at the top of this essay, and then look at those four sets of penis diagrams four column inches below. Both of the shells figured above, and all of those penises, were borne by a single biological species of Pyrgulopsis.
I should conclude this essay, however, emphasizing once again that science is the construction of testable hypotheses about the natural world. Science is not right, it is testable. And over the course of a distinguished career spanning almost 40 years, my buddy Bob rigorously constructed 126 testable hypotheses about the Pyrgulopsis fauna of the great American West. One day, I feel sure, somebody will come behind him and test the 125 that remain. I cannot imagine when, or by whom. Not it.
Notes
[1] Liu, H-P, and R. T. Dillon, Jr. (2021) Resolving the species status of Surprise Valley Pyrg (Pyrgulopsis gibba) and Vineyard Pyrg (Pyrgulopsis vinyardi). Report to Stantec Environmental Consulting. FWGNA Circular 6: 1 – 5. [pdf]
[2] Hershler, R. & H-P. Liu (2017) Annotated Checklist of Freshwater Truncatelloidean Gastropods of the Western United States, with an Illustrated Key to the Genera. US Bureau of Land Management Technical Note 449: 1 – 142.
[3] We always used pine cones when I was growing up, too poor for corn cobs.
[4] Hershler R. 1995. New freshwater snails of the Genus Pyrgulopsis (Rissooidea: Hydrobiidae) from California. The Veliger 38(4): 343-373.
[5] Hershler R. 1998. A systematic review of the hydrobiid snails (Gastropoda: Rissooidea) of the Great Basin, western United States. Part I. Genus Pyrgulopsis. The Veliger 41: 1-132.
[6] The boxed figure of the P. vinyardi penis was scanned from figure 39 of Hershler [5], showing dorsal aspect on the left and ventral aspect on the right. The remainder of the figure, showing penial morphology for three different P. gibba males, was scanned from figure 12 of Hershler [4]. Again, dorsal aspect on the left, ventral on the right. Abbreviations Dg = dorsal gland, Vg = ventral gland, Tg = terminal gland, P = penial filament. The Dg3 region is encircled.
[7] The literature on prezygotic reproductive isolation in Physa is extensive. Here’s a good entry:
- Dillon, R.T., A.R. Wethington, and C. Lydeard (2011) The evolution of reproductive isolation in a simultaneous hermaphrodite, the freshwater snail Physa. BMC Evolutionary Biology 11: 144. [html] [pdf]
[8] Penial morphology of Pyrgulopsis sadai, from Hershler [5], figure 39.
[9] The three CO1 sequences for P. gibba and the one sequence for P. vinyardi were published in four different papers:
- Hershler, R., Frest, T.J., Liu, H.-P., Johannes, E.J. 2003a. Rissooidean snails from the Pit River basin, California. Veliger 46:275-304.
- Hershler, R. and Liu, H.P. (2004) A molecular phylogeny of aquatic gastropods provides a new perspective on biogeographic history of the Snake River Region. Mol. Phylogenet. Evol. 32 (3), 927-937.
- Hershler, R., Liu, H.-P. 2008. Ancient vicariance and recent dispersal of springsnails (Hydrobiidae: Pyrgulopsis) in the Death Valley system, California-Nevada. In: Reheis, M.C., Hershler, R., Miller, D.M., eds. Late Cenozoic drainage history of the southwestern Great Basin and lower Colorado River region: geologic and biotic perspectives. Geological Society of America Special Paper 439:91-101.
- Hershler, R., Liu, H.-P. and Gustafson, D.L. (2008) A second species of Pyrgulopsis (Hydrobiidae) from the Missouri River basin, with molecular evidence supporting faunal origin through Pliocene stream capture across the northern continental divide. J. Molluscan Stud. 74 (4), 403-413.
[10] Ms. Eck actually sent 22 populations for analysis, but population #11 turned out to be a lymnaeid. So just 21 populations of Pyrgulopsis.
[11] In January of 2022 I defined character phase disequilibrium as “any violation of independent assortment between one or more morphological characters and one or more characters of demonstrably genetic origin.” Although CPD can arise from any violation of the assumption of random mating, the most likely explanation in Spring #22 would be reproductive isolation. See:
- What is character phase disequilibrium? [4Jan22]
- Character phase disequilibrium in the Gyraulus of Europe [4Feb22]
[12] Actually I messed up a few, so no count.


Any chance of publishing this finding in a peer reviewed journal? I loved the description of May 2020; that was spot on. So, maybe I am a little lost. I hope I am not being obtuse: are you saying that there are two morphologies across the range of the two taxa, but both morphologies and intermediates are found in this one site? Any clue what is driving the morphology? Is the argument that these are incompletely separated sister taxa, that have come back into contact at site 22?
ReplyDeleteThanks for your kind words. And you've asked some good questions.
DeleteI should have been more clear about the ranges in particular, and will fix that in the main essay as soon as I get off the chat with you. Hershler described P gibba as widespread in the "northwestern Great Basin (CA, NV, OR)." But he considered P vinyardi endemic to "two springs in the Squaw Valley drainage of north central Nevada."
So yes, both morphologies and intermediates are found at Spring #22. I have no hypothesis at all regarding the driver of these morphologies. My hypothesis is, however, that all of these populations are a single polymorphic "taxon," by which I mean biological species.